1.2.2.3. КОРЕНЕВА СИСТЕМА ПОЛЬОВИХ КУЛЬТУР
Загальні питання росту і морфології кореневих систем. Корені є вегетативними органами рослин, за допомогою яких вони вбирають основні елементи мінерального живлення з ґрунту, а також виводять з рослин різні сполуки. Так відбувається обмін речовин у системі ґрунт — рослина. При добре розвиненій кореневій системі рослини ефективно використовують вологу і поживні речовини ґрунту. Дослідження кафедр землеробства і рослинництва Уманської сільськогосподарської академії свідчать, що не лише в орному й підорному шарах, а й глибше (до 3 — 5 м і більше) в ґрунті є достатньо рухомих сполук азоту, калію, кальцію, фосфору та інших макро- й мікроелементів. Тому створенням умов для глибокого проникнення кореневої системи польових культур можна значно зменшити дози внесення добрив без зниження врожаю, що сприяє не лише економії матеріальних засобів, а й, що не менш важливо, отриманню екологічно більш чистої продукції. Корінь, який росте, має так званий кореневий чохлик, під ним безпосередньо розміщується зона клітинного поділу, за нею — зона росту (розтягу) кореня. Клітини в ній витягуються. У них з'являються вакуолі (лат. vacuus — пустий) — порожнини в цитоплазмі клітин, які заповнюються клітинним соком. Завдяки цьому корінь заглиблюється в землю. Якщо цю зону кореня видалити, ріст його донизу припиняється, утворюються бічні корінці. Іноді це роблять штучно з агротехнічною метою на площах насінників люцерни, при пересаджуванні розсади кормових коренеплодів та ін. Вище зони росту розміщена всмоктувальна зона 0,5 — 2 см завдовжки. Вона густо обросла кореневими волосками — тонкими виростками клітин епідермісу 0,2 — 1 см завдовжки. Їх понад 100 штук у всисній зоні. Через них в рослину надходять (всмоктуються) поживні речовини разом з вологою. Це найактивніша (всмоктувальна) зона кореневої системи. Кореневі волоски відмирають через кожні 15 — 20 днів і знову утворюються на ростучих коренях. Ці тонкі корінці — коренева ризосфера — проникають між ґрунтовими частинками і міцно утримують рослину в ґрунті.
Основна маса коренів (65 — 80 %) польових культур на суглинкових ґрунтах Лісостепу й Степу розміщена в шарі ґрунту до 40 см, а з урахуванням довжини коренів найбільше їх (80 - 85 %) у шарі 20 - 60 см.
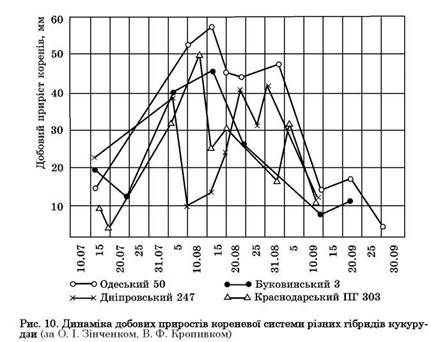
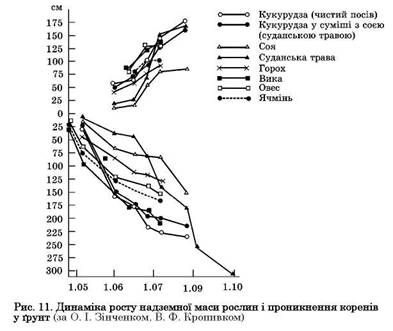
У процесі вегетації рослини ростуть нерівномірно. Залежно від їх біологічних особливостей, а також від умов середовища темпи росту коренів різні, вони неоднаково заглиблюються у ґрунт або майже перестають рости. Під час вегетації польових культур може бути кілька періодів посиленого приросту коренів. Це можна проілюструвати на прикладі динаміки росту коренів різних гібридів кукурудзи (рис. 10). Спостерігається також загальна закономірність вегетації рослин — спочатку посилено ростуть корені, а потім — надземна маса. Початковий ріст коренів сильніший у ранніх ярих — ячменю, вівса, гороху; повільніший — у пізніх ярих (кукурудзи, суданської трави) (рис. 11). Це спостерігається також у проса, сорго та інших просоподібних. Ще повільніше на початку вегетації росте надземна маса цих рослин. Тому ранні ярі досить швидко вкривають площу і стають менш уразливими до бур'янів, ніж пізні ярі, які висівають на 10 — 15 днів пізніше. Пізні ярі засмічуються не лише ранніми бур'янами, а й пізніми та післяжнивними, зокрема мишієм, щирицею, курячим просом (плоскухою) та іншими і потребують старанного догляду на початку вегетації.
|
|
Однією з основних умов посиленого початкового росту коренів є достатнє зволоження орного шару ґрунту. Вміст вологи у суглинкових ґрунтах на початку вегетації має становити 22 — 24 %, супіщаних та піщаних 18 — 20 %. Недостатня зволоженість ґрунту негативно позначається на рості кореневої системи і надземної маси рослин. Існує пряма залежність між зволоженістю ґрунту в період появи сходів і врожайністю культури. За даними О. І. Зінченка, коефіцієнт кореляції (r) цього зв'язку становить для післяукісних 84, для весняних посівів вівса і ячменю 77 — 82, для кукурудзи 67 — 73, післяукісних посівів проса і гречки 84 — 92.
Важливе значення для отримання стабільних урожаїв польових культур має глибина проникнення кореневої системи в ґрунт Польові культури досить відрізняються за цим показником. Корені ячменю, вівса, гороху, сої та інших культур проникають у ґрунт на глибину від 125 - 150 до 200 см; кукурудзи, суданської трави, соняшнику, цукрових буряків, сорго — 250 - 350 до 400 см.
Навіть у періоди несприятливого зволоження в Лісостепу і Степу ґрунт на глибині 1,5 — 2,5 м має достатньо вологи. Це прошарок постійного зволоження, де вологи в материнській породі 22 — 24 %. Корені культур з могутньою кореневою системою (кукурудза, соняшник, люцерна та ін.) досягають цього вологого шару і задовольняють потреби рослин у волозі, якщо її недостатньо в орному і підорному шарах. Тому краще вирощувати культури та їх сорти, які мають добре розвинену кореневу систему.
Глибина проникнення кореневої системи залежить також від ґрунтово-кліматичних умов. За даними Уманської сільськогосподарської академії (Г. І. Мусатов, О. І. Зінченко, В. Ф. Кропивко), в Лісостепу навіть у середньоранніх гібридів кукурудзи коріння проникає на глибину понад 3 м, а в пізньостиглих сортів і гібридів —до 4 м, тоді як на Поліссі корені кукурудзи заглиблюються лише до 2,2 м (В. М. Лебедєв). У Лісостепу й Степу на таку глибину проникають корені пшениці, ячменю, вівса, а в західному Лісостепу, де достатньо вологи в шарі до 100 см, — лише на 1,2 м.
Глибина проникнення коренів у ґрунт залежить не тільки від умов зволоження, а й від механічного та хімічного складу орного й підорного шарів ґрунту. Якщо на глибині 100 — 120 см ґрунт надмірно зволожений, коренева система рослин формується в шарі до 100 см. За таких умов загальний обсяг кореневмісного шару менший і для отримання високого врожаю треба збільшувати норми внесення добрив.
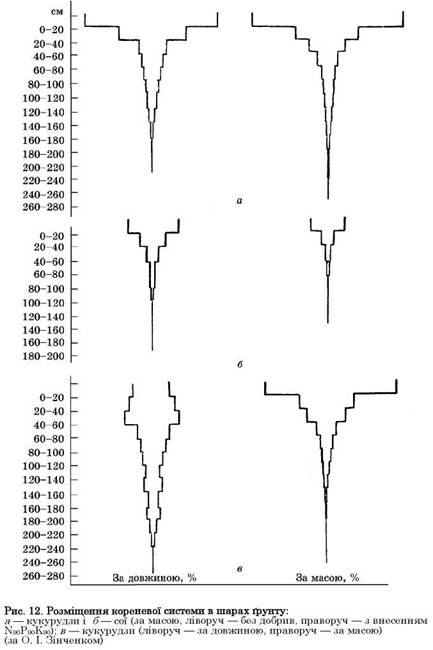
В орному шарі ґрунту розміщується основна маса коренів польових культур. Внесення добрив на посівах злакових сприяє збільшенню маси коренів у верхньому шарі (рис. 12, а). На посівах бобових (в даному разі сої) це не спостерігається, а навпаки, загальна маса коренів може зменшуватись (рис. 12, б). Основна кількість коренів за довжиною (дрібні корінці, так звана ризосфера) розміщується в підорному шарі (рис. 12, в ліворуч),
Важливе значення має коренева система як джерело поживних речовин для наступних культур. Після різних польових культур у ґрунті залишається від 35 — 45 до 70 — 100 ц/га і навіть більше кореневих і стерньових решток. Так, на посівах багаторічних трав щороку залишається в ґрунті 60 — 80 ц/га сухої маси коренів. За 3 — 4 роки використання люцерна залишає 25 — 30 т/га органічної маси, в якій міститься 450 - 500 кг/га азоту, 100 - 120 фосфору, 350 - 400 кг/га калію. Кореневі рештки бобових трав (люцерни, конюшини, вики озимої і ярої, буркуну) містять у два—три рази більше азоту, значно більше фосфору і кальцію, ніж кореневі рештки злакових культур. Навіть 30 — 40 ц/га сухої речовини бобових культур, зокрема вики ярої і мохнатої, залишають 60 — 100 кг/га азоту. Крім того, в процесі вегетації значна його кількість надходить від мінералізації бульбочок та асоціативних азотфіксуючих бактерій, що містяться у ризосфері коренів. Останнє стосується не лише бобових, а й злакових та інших рослин — соняшнику, цукрових і кормових буряків та ін.
Особливості будови кореневої системи польових культур. У польових культур розрізняють два типи кореневої системи — мичкувату (у злакових) і стрижневу (у бобових, хрестоцвітих та інших двосім'ядольних рослин). Є також культури з кореневищним типом кореневої системи, у яких корені є підземними стеблами з міжвузлями, з яких відростають стебла й корені. До них відносяться переважно багаторічні кормові трави — стоколос безостий, лисохвіст лучний, мітлиця. Є також проміжні форми між мичкуватими й кореневищними рослинами (райграс пасовищний, вівсяниця (костриця) червона, тонконіг лучний).
Вегетативним центром у злакових культур є вузол кущення, у бобових, хрестоцвітих та інших — коренева шийка. Процес утворення корінців і надземних пагонів у злакових називають кущенням, у бобових — пагоноутворенням. Кущення і пагоноутворення в однорічних польових культур, крім отавних (відростаючих) кормових трав, відбувається один раз, у багаторічних високоотавних культур —
протягом усього вегетаційного періоду майже безперервно 2 — 5 років і більше.
В однорічних злаків вузол кущення закладається на глибині 1 — 3 см. Між початком його утворення і самим кущенням минає певний час, за який над вузлом кущення закладається певна кількість (4 — 6 і більше) дуже вкорочених міжвузлів і вузлів. Після цього з кожного вузла розвивається один корінець. Коли корінці закріпляться у ґрунті, з кожного вузла розвивається надземний пагін. Пагони розвиваються під кутом до першого — центрального. На кожному з них закладається самостійний вузол кущення, який розвивається у такому порядку, як і головний. Верхня брунька пагона другого порідку під гострим кутом спрямовується догори і виходить на поверхню ґрунту. Період кущення пшениці, жита, ячменю, проса та інших культур досить короткий. Таке кущення називають сим-подіальним. Якби кущення хлібів тривало кілька тижнів, на посівах водночас були б напівстиглі і зелені рослини, тобто хліба з підгоном. Це спостерігається, наприклад, при крайовому ефекті на посівах з широкими технологічними коліями.
Багаторічні злаки, як уже зазначалося, на відміну від однорічних, кущаться безперервно і протягом тривалого періоду. Таке кущення переривається взимку та в бездощові періоди. Кожний пагін багаторічних злаків після нагромадження певної кількості пластичних запасних речовин (білків, жирів, вуглеводів, цукрів) кущиться у свою чергу. Вся маса поживних речовин у результаті фотосинтезу використовується для формування нових пагонів. При цьому зовнішні, більш освітлені, пагони є генеративними, а менш освітлені — вегетативними. Тому травостій багаторічних злаків завжди складається з вегетативних і генеративних пагонів. Для використання на корм більш цінний травостій з більшою кількістю вегетативних пагонів.
На потенційно генеративних пагонах утворюються стебла, і запаси поживних речовин у них використовуються для росту стебла і утворення суцвіть. Після утворення суцвіть та їх цвітіння частину пластичних речовин (яких у пагоні більше, ніж потрібно для наливання і дозрівання зерна) рослина використовує для утворення дочірніх стебел, серед яких також будуть генеративні й вегетативні.
В озимих трав пагони, що утворилися після цвітіння генеративних стебел, виходять у трубку і плодоносять наступної весни. У них цей процес відбувається лише раз протягом року. В ярих високоотав-них злаків він може повторюватись 2 — 3 рази за вегетаційний період.
Головні (генеративні) стебла багаторічних злаків після повного дозрівання насіння видовжуються у нижніх міжвузлях за рахунок інтеркалярного (вставного) росту і вилягають. Зелена маса їх не відмирає до настання морозів. Після цього відмирають плодоносні стебла разом з кореневою системою, листя вегетативних пагонів та вегетативні пагони, у яких накопичилося мало поживних речовин.
Злакові багаторічні трави розвиваються повільніше за бобові. Досить густий травостій їх при звичайних нормах висіву (8 — 10 млн шт. схожих насінин на 1 га) формується лише наступного року після другого скошування, коли після цвітіння почнуть кущитися генеративні пагони.
За достатнього зволоження і посиленого живлення пагоноутво-рення можна посилити і в рік висівання трав. Стимулюють кущення в літньо-осінній період підкошуванням і випасанням худоби на травостоях.
Бобові трави, хрестоцвіті та інші стрижнекореневі рослини розвиваються майже так само, як і багаторічні злаки, але функцію вузла кущення в них виконує коренева шийка (підсім' ядольне коліно). З неї розвивається головний корінь. На кореневій шийці з бруньок (у багаторічних рослин щороку із сплячих бруньок) розвиваються пагони з додатковим корінням. Пагони багаторічних бобових та інших трав щороку відмирають разом з корінцями, поповнюючи запас органічної речовини в ґрунті.
Окрему групу становлять кореневищні рослини, у яких основна маса коріння складається з підземних стебел-кореневищ. Серед зернових, зернобобових культур польової сівозміни їх практично немає, але в кормових травах це досить цінна група рослин. До неї належать пирій повзучий, який за своїми якостями переважає інші злакові трави; стоколос безостий та перехідні форми від кореневищних до нещільнокущових — райграс пасовищний, вівсяниця червона, лисохвіст, мітлиця біла та ін., які мають характерну кореневу систему, що складається з кущів і коротких кореневищ. На них проростають бруньки, з яких догори росте стебельце, а донизу — корінець. Процес такого кущення, або підземного пагоноутворення, так само, як і в нещільнокущових злаків, відбувається безперервно. Від материнської рослини радіально відходять підземні стебла. Кущення злаків відбувається безперервно протягом кількох (іноді багатьох) років. Ці культури продуктивні, щороку накопичують багато органічної маси за рахунок кореневих і стерньових решток (40 — 60 ц/га і більше сухої речовини). З часом органічної маси накопичується багато, внаслідок чого погіршується повітряний режим ґрунту, мінералізація нагромаджених запасів органічної речовини. Подовжують вегетацію кореневищних злаків поліпшенням повітряного режиму ґрунту за допомогою розпушування дисковим знаряддям, голчастими боронами.
Аналогічний повзучий тип пагоноутворення характерний і для деяких бобових трав. Однак стебло у них розміщується на поверхні ґрунту, наприклад у конюшини білої. Такі рослини відростають добре після спасування.
Наливання і достигання зерна. Після закінчення цвітіння починається утворення зерна. При наливанні зернівка наповнюється вуглеводами, цукрами, білками і спершу має рідку консистенцію, так звану молочну стиглість. У цю фазу стиглості уже практично повністю формується зародок, зерно в цій фазі стиглості вже може проростати. При достиганні в ендоспермі або в сім'ядолях бобових та інших двосім'ядольних рослин збільшується вміст сухих речовин, зерно набуває густої консистенції, настає молочно-воскова фаза достигання, яка змінюється восковою, після воскової — фаза повної стиглості зерна. На початку фази зерно має підвищену вологість (35 — 40 %), а після повного достигання зневоднюється до залишкової вологості 12 — 14 %. Таке зерно можна зберігати протягом тривалого періоду. На кормові цілі можна використовувати зерно вологістю 35 — 40 % у спеціальних герметичних місткостях. Таким способом зберігають вологе ціле і подрібнене зерно кукурудзи й сорго. На корм великій рогатій худобі зерно кукурудзи подрібнюють разом з качанами. Якщо подрібнених стрижнів качанів у заготовленій масі небагато, то її можна згодувати і свиням.
Зерно кукурудзи у фазі молочної стиглості також використовують для харчових цілей. Дуже цінним харчовим продуктом є заморожені качани. В такій фазі стиглості консервують і зерно гороху.
Описані вище загальні екологічні та біологічні особливості польових рослин взаємопов'язані. Сорти й гібриди культур розрізняються насамперед екологічними особливостями: висотою стеблостою, будовою листя, кущистістю, стійкістю проти ураження шкідниками і хворобами, морозо- і зимостійкістю, посухостійкістю та ін. Отже, сорт — це екотип, який набуває певних екологічних ознак в результаті селекційної роботи, він же є біотипом, оскільки може відрізнятися від іншого тривалістю проходження фаз вегетації або міжфазними періодами, температурою проростання насіння, відношенням до умов проходження стадії яровизації, врожайністю, якістю врожаю тощо. Так, можуть бути біотипи озимих пшениці, жита, ячменю з тривалим періодом стадії яровизації і з коротким, що, в свою чергу, є важливим при визначенні строків осінньої сівби. Тому екологічні і біологічні особливості треба враховувати при розробці сортової технології вирощування високих врожаїв польових культур.