2.3. Водний режим і продуктивність рослин при зрошенні
Вода, будучи однією із важливіших умов життя рослин, по закону взаємодії факторів, необхідна в оптимальних кількостях також для більш повного використання сільськогосподарськими культурами всіх інших факторів формування врожаю.
У районах зрошуваного землеробства багато світла і тепла. Ґрунти мають порівняно хорошу родючість. Проте повністю використати ці сприятливі природні умови рослини не можуть внаслідок недостатнього природного зволоження. Зрошення усуває недостачу вологи, її обмежуюча дія на продукційні процеси припиняється, і урожайність сільськогосподарських культур збільшується в декілька разів.
Участь води в процесах росту і розвитку рослин. Вода прямо чи побічно бере участь в усіх процесах, які відбуваються в рослинах Разом з водою з ґрунту надходять до рослин поживні речовини і транспортуються до листків та інших органів; пластичні речовини, з яких утворюються нові живі клітини, пересуваються з листків до ростучих органів також за допомогою води.
Вода необхідна і як середовище, в якому відбуваються всі біохімічні перетворення. Від ступеня насичення тканин рослини водою залежить швидкість і напрям цих перетворень.
Без участі води неможливий фотосинтез, в результаті якого утворюється органічна речовина. Фізіологи відносять також воду до тих речовин, елементи яких витрачаються і знову синтезуються в обміні речовин. Виникає поняття про метаболічну воду.
Вода безпосередньо впливає на ріст рослин К.А. Тімірязєв вважав, що ріст рослин та їх окремих органів зводиться в кінцевому результаті до засвоєння води. Зв'язок між водозабезпеченістю і ростом виражається у збільшенні висоти рослин, кількості та розмірів листків, гілок, у прискоренні росту плодових та інших органів При підвищенні вологості ґрунту поліпшується ріст кореневої системи і збільшується її маса.
За нашим обліком, проведеним в учбовому господарстві «Приозерне» Херсонського СГІ, маса кореневої системи кукурудзи в шарі 0‑40 см під впливом зрошення збільшилася в два рази; маса коріння люцерни в тому ж шарі становила: без поливів 8,8 т, при поливі прісними водами – 14,1 і при поливі стічними водами – 17,3 т/га.
Як відомо, продукційні процеси здійснюються, головним чином, у листках При зрошенні їх площа нерідко збільшується в декілька разів.
В.С. Снєговий (1967) відзначив збільшення площі листків однієї рослини сої у зоні дії Інгулецької зрошувальної системи від 1505 на контролі (без зрошення) до 4530 см2 у кращому варіанті поливу.
Вологозабезпеченість рослин суттєво впливає на умови їх повітряного живлення. Від насичення тканин листків водою залежить ступінь відкриття продихів, через які надходить вуглекислий газ і відбувається газообмін між листками і повітрям.
Поряд із збільшенням вмісту в листках води і поліпшенням їх повітряного живлення поливи створюють сприятливі умови для підвищення в листках вмісту хлорофілу. Так, на посівах кукурудзи у фазі молочної стиглості вміст хлорофілу у листках без поливу становив 4,8 мг, при зрошенні прісними водами – 5,5 і при поливі стічними – 8,1 мг на 1 кг сухої речовини. Все це сприяє збільшенню інтенсивності фотосинтезу, а також його продуктивності.
Узагальнюючи дослідження на півдні України, О.О. Собко (1975) відзначає таке збільшення продуктивності фотосинтезу озимої пшениці: у період колосіння – цвітіння від 4,41 г/м2 на добу без поливу до 6,10 в умовах оптимального зрошення; у період молочно-воскової стиглості відповідно 2,51 і 6,32 г/м2 за добу.
Збільшення площі листків і вмісту в них хлорофілу при зрошенні є важливою умовою ефективного використання рослинами сонячної енергії.
О.О. Собко (1975) наводить такі дані про засвоєння посівами озимої пшениці, яка знаходиться у фазі цвітіння, фотосинтетично активної радіації (ФАР): сорт Безоста 1 без поливу – 82,4 тис. кДж/га за 1 хв., в умовах оптимального поливного режиму – 195,2; сорт Кавказ відповідно 100 і 215,2.
При невеликій втраті води листками активність фотосинтезу в ряді рослин зменшується незначно, але помітно затримуються відтік асимілятів з листків, ріст і розвиток. При великих втратах води (понад 15‑20%) депресія фотосинтезу і порушення інших фізіологічних процесів у рослині різко зростають.
В.А. Бриліант (1970) вважає, що зміна активності фотосинтезу при зниженні обводненості рослинних клітин залежить від виду і фаз розвитку рослин, ступеня і способу зневоднення, умов освітлення та ін. При зневодненні порушується нормальний стан колоїдів цитоплазми, що зумовлює хід фотохімічних і темпових реакцій, фотосинтезу.
Дослідження показують також, що при недостатньому водопостачанні змінюється склад первинних продуктів фотосинтезу, зокрема затримується утворення фосфорних ефірів цукру. Частка низькомолекулярних сполук у загальній сумі асимілятів у посуху збільшується.
При втраті води листками посилюється дихання рослинної тканини, а при глибокому зневодненні погіршується загальний стан біоколоїдів клітини.
Водний режим впливає на розвиток рослин. В одних випадках настання фаз при оптимальному водному режимі прискорюється (з'явлення сходів, кущіння тощо), розвиток репродуктивних органів нерідко затримується. Це необхідно враховувати при вирощуванні сільськогосподарських культур.
Кількість і стан води в рослинах. Рослина більше ніж наполовину складається з води. У листках її більше, ніж у корінні. За даними М.А. Максимова (1971), листки містять 80‑90% води, а коріння – 70‑80% до сирої маси.
Вода в рослинах міститься в стані, що різниться за ступенем рухомості окремих її фракцій, які, однак, знаходяться в тісній взаємодії між собою. Вода разом з розчиненими в ній хімічними сполуками заповнює судинну систему рослин, вакуолі клітин, вступає в рухливий зв'язок з білками та іншими складовими частинами клітин. Цитоплазма включає воду як один із своїх компонентів (75‑85% цитоплазми; Алексєєв А.М., 1969). Вона, як і інші органели клітин, діє на молекули води і обмежує їх рухливість (упорядкований або структурований стан води). Чим більша структурованість води, тим менша її рухливість.
В. Лархер (1978) розрізняє такі форми води в клітині: конституційна, або хімічно зв'язана; гідротаційна, що утворює шари навколо іонів розчинених органічних речовин і макромолекул і проникає в проміжки між ультраструктурами цитоплазми і клітинної стінки; резервна, що заповнює водозбірні порожнини і вакуолі; інтерстиційна, що виконує транспортні функції у міжклітинниках і в провідних шляхах – судинах і ситоподібних трубках.
Гідратаційна вода в цитоплазмі й клітинній стінці є здебільшого капілярнозв'язаною. До гідратаційної належить і колоїдно зв'язана, яка становить 5‑10% внутрішньоклітинної води. Невелика втрата цієї води може призвести до пошкодження клітинних структур і до загибелі клітин.
Резервна вода, що заповнює резервуари розчинів, рухлива, але не цілком вільна, вона зв'язана осмотично з розчиненими речовинами, кількість і стан яких у клітині регулюється шляхом полімеризації і гідролізу.
Багато авторів відмічають з'єднання кількості найважче віднятої води з клітин із вмістом білків Частина води випаровується з поверхні рослинних клітин у міжклітинники і знаходиться у пароподібному стані.
Узагальнюючи дані багатьох досліджень, М.С. Петинов (1962) відзначає, що визначені фракції води зв'язані з рядом фізіологічних процесів – транспірацією, фотосинтезом, диханням, ростом.
При недостатньому водопостачанні насамперед зменшується вміст у рослині найбільш рухомої (менш упорядкованої) води, тим часом як наявність визначеної її кількості й активність в органах рослин дуже важливі для їх нормальної життєдіяльності і високої продуктивності.
У житті рослин важливу роль відіграє і більш зв'язана вода. Зневоднення колоїдів цитоплазми призводить до порушення біологічних процесів. Особливо небезпечне швидке зневоднення, при якому можуть відбуватися необоротні порушення структури найважливіших складових частин живої клітини.
Протидія, або опір, відняттю води з рослинних клітин називається їх водостримуючою властивістю. Опір пов'язаний з розривом водневих зв'язків і електростатичних сил притягання між молекулами. Водостримуюча властивість зростає в міру зменшення води в клітинах.
Втрата води рослинами – це не тільки фізичне явище. При зневодненні відбувається перебудова білків і всього біокомплексу рослинних тканин, що конкретно впливає на водний режим рослин.
При недостатній вологості ґрунту порушується водний режим як у наземній частині рослин, так і в коріннях. У них різко збільшується відносна кількість більш зв'язаної (упорядкованої, структурованої) води, послаблюється подача соку, значно знижується кількість фосфору в ній. При нормальному водопостачанні в коріннях синтезуються складні органічні сполуки фосфору. При нестачі ж води кількість їх зменшується, так само як і вміст неорганічного фосфору.
При нормальному водопостачанні переважають процеси синтезу органічної речовини в рослині, а при зневодненні посилюється ферментативний розпад складних сполук клітини.
Зрошення сприяє відновленню нормальної обводненості всіх органів рослини, внаслідок чого створюються сприятливі умови для синтезу. Синтез органічної речовини при цьому переважає над розпадом, що призводить до швидкого нагромадження сухих речовин у рослинах.
Транспірація та її значення для рослин. З усієї води, що поглинається корінням з ґрунту, рослина засвоює незначну кількість – 0,15‑0,20% і лише іноді більше. Залишок води випаровується з поверхні листків та інших надземних органів рослин. Цей процес називають транспірацією.
Завдяки транспірації відбувається безперервний потік вологи з ґрунту до кореневої системи, із коренів у стебла, гілки, плодові органи і листки, а з них в атмосферу Треба враховувати, що механізм надходження води з ґрунту до кореневої системи складний і залежить від стану рослини і ґрунтових умов.
Втрату води внаслідок транспірації не можна вважати некорисною. Завдяки їй полегшується надходження води разом з мінеральними солями до листків та інших органів рослин, що забезпечує нормальне живлення.
При транспірації відбувається охолодження листків, що запобігає їх перегріванню. Спостереження показує, що температура листків вище 35°С несприятлива для фізіологічних процесів. Така температура повітря – звичайне явище в південних районах у найбільш жаркий час дня. Внаслідок транспірації температура листків буває на 3‑8° нижче навколишнього повітря. Про зниження температури листків сої при поливі свідчать дані таблиці 4. Температура листків при поливі знижується внаслідок витрати тепла на транспірацію і зменшення випромінювання його ґрунтом.
4. Вплив поливу на транспірацію і температуру листків сої
(Снєговий В. С., 1967)
|
Дата спостереження |
Варіант |
Вологість ґрунтів у шарі 0-70 см,% НВ |
Інтенсивність транспірації, мг/хв/см2 |
Температура поверхні листків,°С |
Температура поверхні ґрунту,°С |
|
13 липня |
Без поливу |
63,5 |
0,144 |
34,4 |
46,0 |
|
13 липня |
Полив |
82,0 |
0,465 |
31,3 |
26,0 |
|
17 липня |
Без поливу |
58,5 |
0,192 |
35,4 |
49,5 |
|
17 липня |
Полив |
72,0 |
0,616 |
33,2 |
32,0 |
При добрій вологозабезпеченості рослин транспірація посилюється. При зрошенні підтримується певний запас води в ґрунті і певний рівень транспірації. Однак надмірне посилення транспірації небажане. Воно призводить до непродуктивних витрат води на одиницю врожаю. Для регулювання цього процесу потрібно знати, які умови визначають кількість води, що випаровується рослинами.
Величина транспірації значною мірою визначається біологічними особливостями культури і сорту. Невисоким транспіраційним коефіцієнтом, що показує витрати води в грамах при утворенні 1 г сухої речовини, відрізняються кукурудза, просо; у пшениці, ячменю та інших зернових колосових він становить у середньому 400‑500 одиниць; у люцерни, картоплі – 600‑700. Транспіраційні коефіцієнти неоднакові також у різних сортів однієї і тієї самої культури.
Умови зростання визначають рівень транспірації не в меншій мірі, ніж біологія рослин. Відзначимо, що при високій температурі і сухості повітря відносна величина транспірації збільшується.
За даними Безенчуцької дослідної станції, транспіраційний коефіцієнт пшениці в сухий і жаркий рік становив 628, а у вологий – 316, у ячменю відповідно 618 і 288, у кукурудзи – 437 і 160. Ці величини одержані при однаковій вологості ґрунту, різними були тільки умови повітряного середовища. При підвищенні температури сухість повітря зростає. Ці умови пов'язані між собою.
На величину транспірації впливають умови освітлення і швидкість вітру. При недостачі світла затримується нагромадження сухої речовини, а листки мають крупні тонкостінні клітини, погано захищені від випаровування, і транспіраційний коефіцієнт збільшується 3 підвищенням швидкості вітру транспірація також збільшується.
Доречно відзначити загальновідомий факт, що при зрошенні вологість повітря в приземних шарах збільшується, а температура в денні години знижується. Отже, умови повітряного середовища створюються більш сприятливо для обмеження величини транспірації.
На транспірацію в значній мірі впливають грантові умови. Чим вища вологість ґрунту, тим вільніше він віддає свою воду росли нам, і коефіцієнт транспірації підвищується. Ця закономірність відмічена ще в роботах академіка Д.М. Прянишникова. Наприклад, у його дослідах транспіраційний коефіцієнт вівса при вологості ґрунту 40, 60 і 80% становив відповідно 402, 483 і 505.
Транспіраційний коефіцієнт збільшується повільніше, ніж підвищується вологість ґрунту.
При поліпшенні ґрунтового живлення рослин вода використовується більш економне, і транспіраційний коефіцієнт зменшується. Наприклад, у тому ж досліді Д. М. Прянишникова при внесенні добрив транспіраційний коефіцієнт при вологості ґрунту 60% ПВ зменшився від 505 до 409 (табл. 5)
5. Величина транспіраційного коефіцієнта вівса
(за даними вегетаційного досліду)
|
Варіант |
Вологість ґрунту, % ПВ | ||
|
40 |
60 |
80 | |
|
Без добрив |
402 |
483 |
505 |
|
З добривами |
334 |
372 |
409 |
Знання цих закономірностей зобов'язує працівників зрошуваного землеробства поєднувати зрошення з правильним використанням добрив і застосуванням інших прийомів, які поліпшують ґрунтове живлення рослин. Це забезпечує більш економне і раціональне використання зрошувальної води у виробничих умовах.
Вологолюбність і посухостійкість рослин. Різні рослини та їх сорти відрізняються неоднаковою потребою в воді і по-різному переносять її нестачу. Відносно високої вологості ґрунту потребують рис, овочеві культури, лучні трави. Для скоростиглих кормових культур і хлібів потрібний порівняно менше зволожений ґрунт.
Посухостійкі культури можуть добре використовувати додаткове зволоження і давати при цьому високі врожаї. Однак збільшення врожаю від зрошення значно коливається залежно від культури і сорту.
Для умов зрошення необхідно підбирати не тільки водовимогливі та цінні в господарському відношенні культури, але і створювати спеціальні сорти, пристосовані до цих умов. Бажано поєднувати в таких сортів властивість високоефективно використовувати воду з жаростійкістю і посухостійкістю. Насамперед у нових сортів важлива стійкість проти повітряної посухи, оскільки зрошення розвивається в зоні, де цей вид посухи дуже поширений і для якої характерна висока температура. Велике значення має також стійкість проти можливого неповного водопостачання у між-поливні періоди, що не виключено в господарських умовах.
Посухостійкість більше визначається потужністю кореневої системи рослин, а не витратою води на транспірацію. Чим глибше коріння проникає в ґрунт, тим надійніше забезпечується водопостачання рослин за рахунок глибинних запасів вологи. Крім того, посухостійкі рослини мають захисні пристрої від надмірного витрачання води. Тому не можна пов'язувати посухостійкість рослин тільки з величиною транспіраційного коефіцієнта. Цей мінливий показник має лише відносне значення для характеристики рослини і не вичерпує його відношення до вологи.
За дослідженнями М.А. Максимова (1971), посухостійкі рослини характеризуються також дрібноклітинністю будови, густою сіткою провідних пучків, великою кількістю продихів на одиницю поверхні листків і малими їх розмірами. Важливіші ділянки тканин листка в посухостійких рослин захищені від перегрівання і зневоднення відкладаннями крохмалю. Однією з найважливіших особливостей посухостійких рослин є їх властивість переносити з найменшою шкодою тимчасове зневоднення тканин.
До найбільш посухостійких культур належать просо, сорго, суданська трава, кукурудза, соняшник; середньо- і слабостійкі проти посухи пшениця, цукрові буряки, люцерна, соя, рицина, горох, картопля, помідори; найменш стійкі – рис, капуста, огірки.
Критичні періоди споживання води рослинами. Вода необхідна рослинам починаючи з набубнявіння і проростання насіння до достигання плодів. На початку вегетаційного періоду рослини споживають відносно невелику кількість вологи, потім споживання зростає і досягає максимуму в період запліднення, зав'язування і активного росту плодів або в строки, близькі до цих фаз, а до кінця вегетації, як правило, зменшується.
Нестача вологи в усі періоди розвитку сільськогосподарських культур знижує їх продуктивність. Проте нестача води призводить до різких результатів залежно від фази розвитку рослин. У їх житті виділяють порівняно короткі періоди, протягом яких при недостатній кількості вологи особливо різко знижується урожай, а достатнє водопостачання забезпечує високий його рівень.
Такі періоди в рослин вперше виявив російський учений П.І. Броунов і назвав їх критичними.
Посуха особливо шкідливо впливає на молоді органи, що знаходяться у фазі активного росту. Тому критичний період для рослин звичайно припадає на час, коли початкову фазу росту і розвитку проходять органи, що визначають урожай. У багатьох сільськогосподарських культур це репродуктивні органи – плоди і насіння. Природно, що критичні періоди нерідко спостерігаються перед цвітінням або відразу ж після нього. У зв'язку з цим рослини пристосовувалися до того, щоб забезпечувати краще водопостачання молодих репродуктивних органів. Так, глибоке проникнення до цього часу коренів у ґрунт збільшує обсяг доступної вологи. У деяких рослин при з'явленні перших зав'язей листки скорочують випаровування вологи, внаслідок чого зростає її приплив до плодових гілок і молодих зав'язей.
|
За Ф. Д. Сказкіним, критичні періоди настають у такі фази: | ||
|
Озима пшениця, ячмінь, овес |
– |
вихід у трубку-колосіння |
|
Просовидні (сорго, просо) |
– |
викидання волоті |
|
Кукурудза |
– |
цвітіння-молочна стиглість |
|
Бобові, гречка, гірчиця |
– |
цвітіння |
|
Соняшник |
– |
утворення кошиків-цвітіння |
|
Баштанні |
– |
цвітіння-достигання |
|
Буряки на насіння |
– |
стрілкування-цвітіння |
|
Картопля |
– |
цвітіння-формування бульб |
|
Помідори |
– |
цвітіння-формування плодів |
Причини різкого зниження врожаю при нестачі вологи в критичні періоди у різних рослин можуть бути різними. Так, за даними Ф.Д. Сказкіна (1961), у ярих хлібів у цей період може пошкоджуватися пилок у пиляках, що призводить до череззерниці та з'явлення пустих колосків. У рослин, урожай яких становлять вегетативні органи, критичною з виробничої точки зору є фаза активного росту таких органів. Для цукрових буряків це період активного росту коренеплодів, для картоплі – бульб і т. д.
Поняття критичного періоду не можна пояснювати спрощено, пов'язуючи величину врожаю тільки з умовами водопостачання в цей час. Вона визначається також, хоч і в меншій мірі, умовами до- і післякритичного періоду. Крім цього, в критичний період рослина найбільш чутлива не тільки до нестачі вологи, а й до надходження поживних речовин.
Виробниче завдання, що випливає з цих закономірностей, полягає в тому, щоб за допомогою зрошення безперебійно забезпечувати рослини вологою під час вегетації згідно з їх потребами за періодами розвитку. При цьому необхідно знати строки проходження критичних періодів і в цей час по можливості повністю задовольняти потребу рослин у воді. Треба також враховувати, що рівень оптимальної передполивної вологості в критичні періоди звичайно вищий, ніж у інший період вегетації.
Важливо враховувати також, що критичний період не завжди збігається з періодом максимального споживання води рослинами. Останній визначається не тільки біологічним станом рослин, але й метеорологічними умовами: високою температурою, сухістю повітря, суховіями та ін. У таких умовах не можна обмежувати споживання води рослинами, оскільки це призвело б до погіршення їх фізіологічного стану і зниження продуктивності.
Вода і водні властивості ґрунту. Волога дощу, талих і іригаційних вод поглинається поверхнею ґрунту і просочується вглиб.
У цьому процесі, що називається інфільтрацією, розрізняють стадії вбирання і фільтрації. При вбиранні вода проникає в ґрунт через поверхню і поглинається нею, при фільтрації здійснюється подальше пересування і перерозподіл увібраної вологи в ґрунті. Якщо ґрунт має дрібні капілярні пори, пересування води у ньому надто утруднено. Вода, що заповнює некапілярні пори, легко і швидко пересувається вниз під дією сили тяжіння. Швидкість пересування вологи виражається коефіцієнтом фільтрації: на середніх і легких суглинках – 0,05‑0,10 м/добу, супісках – 0,10‑0,50, лесу – 0,25‑0,50, піску пилуватому – 0,5‑1,0, піску дрібнозернистому – 1,0‑5,0.
Стан, коли всі пори ґрунту заповнені водою, відповідає повній вологоємності (ПВ). Повна вологоємність вираховується найбільшою кількістю води, яку може вмістити ґрунт, вираженою в процентах маси його абсолютно сухої речовини або в інших одиницях (кубічних метрах, міліметрах). Зволоження ґрунту до 100% ПВ – явище рідкісне і нетривале в посушливих районах. Воно спостерігається протягом короткого часу після рясних дощів, танення снігу, поливу. Як тільки вода, що заповнює крупні пори, стече вглиб, залишається її запас, надійно затриманий верхнім шаром ґрунту.
Найбільша кількість води, яка надійно затримується ґрунтом, відповідає найменшій вологоємності, синонім – польова вологоємність.
При глибокому заляганні підґрунтових вод ґрунт кореневмісного шару, зволожений до найменшої вологоємності, не містить воду, здатну пересуватися під дією сили тяжіння.
При близькому заляганні підґрунтових вод, внаслідок їх підпору, вологість ґрунту підвищується і відповідає капілярній вологоємності (KB), (рис. 2).
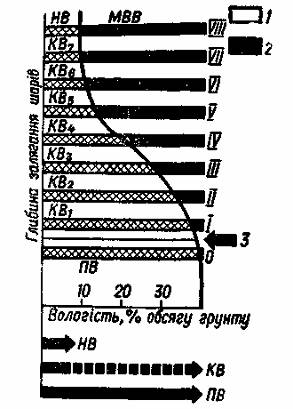
Рис. 2. Співвідношення між повною (ПВ), капілярною (КВ) і найменшою
вологоємністю (НВ) та максимальною водовіддачею (МВВ):
1 – вода, що утримується в ґрунті;
2 – вода, віддана ґрунтом;
3 – початковий рівень підгрунтових вод.
Повна і найменша вологоємність – важливі для зрошуваного землеробства показники властивостей ґрунту. Повна вологоємність показує, яку найбільшу кількість води може прийняти ґрунт при зрошенні; найменша вологоємність характеризує найбільшу кількість води, яку ґрунт може надовго утримувати після рясного поливу. При зволожувальному зрошенні частіше доцільно доводити вологість ґрунту саме до цього стану.
Властивість ґрунту віддавати частину води шляхом стікання її під дією сили тяжіння називається водовіддачею (ВВ). Найбільші показники водовіддачі відмічаються при глибокому заляганні підґрунтових вод. Це максимальна водовіддача. Величину водовіддачі визначають за різницею між повною і найменшою вологоємністю.
З поглибленням шару ґрунту водовіддача звичайно зменшується. Залежно від характеру ґрунтових пор, механічного складу ґрунту його водопідйомна властивість різна. Чим менші й одно рідніші ґрунтові пори, тим сильніше виражена ця властивість. Водопідйомна властивість невелика в структурних і піщаних ґрунтах (З0‑60 см), але значна (більше 2 м) у безструктурних глинистих ґрунтах.
Про величину водопідйомної властивості ґрунту можна судити по висоті шару з підвищеною вологістю, який знаходиться над рівнем підґрунтових вод. Тут вологість ґрунту підвищується внаслідок підпору підґрунтових вод і підняття води по капілярах. Тому цей шар дістав назву капілярної зони. У межах її вологість тим більша, чим ближче рівень підгрунтових вод.
Завдяки водопідйомній властивості ґрунту рослини можуть використовувати підгрунтові води (при визначених умовах їх залягання). Надто близький рівень підгрунтових вод може бути причиною заболочування і засолення ґрунту.
До важливих водних властивостей ґрунту належить також його здатність зв'язувати рідку і пароподібну воду. Дипольні молекули води, тобто молекули, які мають, незважаючи на свою електронейтральність, позитивний і негативний полюси, можуть зв'язуватися одна з іншою полюсами протилежного знаку, а також з іншими тілами, що мають електричний заряд. Зв'язана ґрунтовою часткою вода відрізняється орієнтованістю: всі дипольні молекули її повернені до частки визначеними полюсами.
Деяка частина рідкої води міцно зв'язується з ґрунтом. При цьому виділяється теплота змочування (до 41‑62 Дж/г ґрунту). Це показує, що міцно зв'язані молекули води, втрачаючи кінетичну енергію, переходять у практично непорушний стан. При цьому густина води різко зростає (іноді до 1,7) і знижується її здатність розчиняти солі.
Крім міцнозв'язаної води, ґрунтові частки здатні утримувати ще деяку кількість рихлозв'язаної води, яка менше відрізняється за своїми властивостями від звичайної. Зв'язана ґрунтом вода створює водяну оболонку її часток, так звану плівкову воду.
Здатність ґрунту поглинати і зв'язувати молекули пароподібної води називається її гігроскопічністю. Найбільша кількість пароподібної води, яку здатний поглинати ґрунт, відповідає її максимальній гігроскопічності (МГ). При зволоженні, що відповідає максимальній гігроскопічності, ґрунт містить міцнозв'язану і частково рихлозв'язану воду, причому міцнозв'язана вода становить 60‑80% МГ.
Водні властивості ґрунту залежать від його механічного складу, ступеня гумусності, складу поглинутих основ, структурності будови та інших показників. Чим краща ґрунтова структура, тим більша водопроникність. Тимчасово водопроникність ґрунту може бути поліпшена обробітком ґрунту, насамперед глибоким розпушенням.
Із збільшенням вмісту мулистих часток у ґрунті та ступеня його гумусності зростають максимальна гігроскопічність і вологоємність ґрунту.
Споживання води рослинами і біологічна оцінка різних ступенів зволоження ґрунту. Надходження води до рослин пов'язане, з одного боку, з властивістю ґрунтової вологи пересуватися до кореневих волосків, які висушують ґрунт, з другого – з властивістю коренів рухатися до нових джерел ґрунтової вологи під час росту. Перша умова особливо важлива для нормальної життєдіяльності рослин. Вода ґрунту знаходиться в різному стані, що визначає ступінь доступності її для використання рослинами.
Основи класифікації ґрунтової вологи з оцінкою її рухливості дав А.Ф. Лебедев (1912). А.А. Роде (1965) пропонує розрізняти такі агрономічне важливі категорії, форми і види ґрунтової води.
Вільна вода: вільна гравітаційна – пересувається тільки під впливом сили тяжіння; підперта гравітаційна – утримується капілярними силами; підвішена – підтримується капілярними і сорбційними силами.
Міцнозв'язана вода утворює на поверхні часток ґрунту тонку плівку, товщина якої вимірюється двома – трьома діаметрами молекул води За своїми якостями близька до твердого тіла, має високу густину.
Рихлозв'язана вода утворює навколо ґрунтових часток плівку, товщина якої може досягати десятків діаметрів молекул води. Густина не перевищує густини звичайної рідкої води. Внутрішні шари мають зрушувальну міцність.
Пароподібна вода знаходиться в ґрунтовому повітрі у вигляді водяного пару. Рухається вона в бік більш низької пружності. Може переходити в рідку форму.
Вільна волога найрухливіша і найдоступніша для рослин. Однак вільна гравітаційна волога в ґрунті носить тимчасовий характер внаслідок руху в глибинні шари ґрунту, тоді як капілярна більш стійка і є основним джерелом водопостачання рослин.
Пори шириною до 60 мкм затримують воду завдяки капілярним силам (Лархер В., 1978). Така вода утримується ґрунтом незалежно від глибини підґрунтових вод, тобто й при відсутності підпору. У цьому випадку вона називається капілярно підвішеною.
У найбільш крупних порах, шириною понад 60 мкм, слабше проявляється капілярний підйом води і добре виражене стікання під дією сили тяжіння. Стікання припиняється при підпорі підґрунтових вод. У крупних порах над ґрунтовою водою міститься підперта гравітаційна вода. Вона вільно використовується рослинами, якщо їх коріння проникає у капілярну зону.
Рухливість капілярної води залежить від величини пор. У дуже дрібних порах, наприклад, у надмірно ущільненому ґрунті, вся вода може бути в зв'язаному стані.
У підвішеному стані, крім капілярної, знаходиться плівкова вода, що обволікає ґрунтові частки або вистилає стінки пор (міцно- і рихлозв'язана вода), а також сорбційно-замкнена вільна і в пустих порах, пароподібна волога.
Всі форми підвішеної вологи становлять її запас, який відповідає найменшій вологоємкості.
Частина підвішеної води здатна до пересування в бік більшого висушення ґрунту. Але цей рух затримується при зменшенні вологості ґрунту до визначеної межі, яка називається вологістю розриву капілярного зв'язку (ВРК). Ця межа має важливе біологічне значення, оскільки вказує на погіршення умов водопостачання рослин. Вона відповідає приблизно 60‑70% НВ Найменші показники межі відмічаються на легких ґрунтах.
За спостереженнями С.І. Долгова, до ВРК близька вологість уповільнення росту рослин. Цей показник названий вологістю гальмування (ВГ) Він більш точно, ніж ВРК, відображає індивідуальні особливості водопостачання окремих культур, їх сортів або гібридів.
При вологості ґрунту менше ВРК переважає плівкова волога. А.А. Роде (1965) відзначає, що не менше двох третин всієї тієї кількості вологи, яка міститься в ґрунті при вологості, що дорівнює НВ, знаходиться у формі плівки. Це міцно- і рихлозв'язана вода. Яка їх доступність для рослин?
Міцнозв'язана вода абсолютно недоступна для рослин. Усмоктувальна сила гігроскопічне зв'язаної води, за В. Лархером, 5 млн/Па. Це мертвий запас вологи, при якому рослини гинуть. Після зменшення вологості ґрунту до мертвого запасу поливи не відновлюють життєдіяльність рослин.
Внутрішні шари рихлозв'язаної води, ближче до поверхні ґрунтової частки, практично нерухомі, утримуються силами в 1,5 млн/Па і більше. Ця вода важкодоступна для рослин.
Кореневий волосок здатний поглинути молекули такої води, які знаходяться з ним у безпосередньому зіткненні, але водоспоживання цим обмежується, оскільки волога не пересувається до точок висушування. При її наявності рослина знаходиться в стані стійкого в'янення, на грані загибелі. Всмоктувальна сила в 1,5 млн/Па відповідає вологості в'янення рослин і границі продуктивної вологи.
Периферійна частина плівкової води, найменше зв'язана сорбційними силами, може переміщуватися і доступна рослинам, але не забезпечує їх нормальної продуктивності (рис. 3).
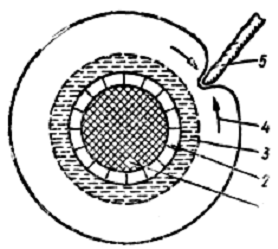
Рис. 3. Схема використання кореневим волоском рихло-зв'язаної води:
1 - ґрунтова частка;
2 - міцно-зв'язана вода;
3 - внутрішні шари рихлозв'язаної води, які відзначаються зрошувальною міцністю;
4 - периферійні, рухливі шари рихлозв'язаної води;
5 - кореневий волосок
Пароподібна вода ґрунту безпосередньо не використовується рослинами, але повне насичення нею ґрунтового повітря необхідне для нормальної життєдіяльності кореневої системи. Такі умови створюються при вологості, що перевищує нижню границю продуктивної вологи. При визначених умовах пароподібна вода може компенсуватися, переходячи в рідкий стан.
Згідно з викладеними вище положеннями С.І. Долгов (1957) дає наступну біологічну оцінку різних ступенів зволоження ґрунту, виділяючи шість ступенів зволоження: волога, недоступна для рослин (міцнозв'язана); недостатня, важкодоступна (інтервал від ВЗ – вологості в'янення до «мертвого запасу»); доступна, але заниженої продуктивності (переважають периферійні шари рихлозв'язаної вологи); доступна, нормальної продуктивності – інтервал від ВРК до НВ; висока легкодоступна; надмірна (нестача повітря).
Поливами підтримується четвертий ступінь зволоження. Він відповідає діапазону оптимальної вологи для рослин: його верхня межа НВ, нижня – ВРК або ВГ для окремих культур. Таким чином, діапазон оптимальної вологи ДОВ = НВ‑ВРК, або, точніше, ДОВ = НВ‑ВГ.
У сільськогосподарській практиці часто визначається діапазон продуктивної вологи (ДПВ), його межі НВ і ВЗ, отже, ДПВ=НВ‑ВЗ. За термінологією А. А. Роде, це діапазон активної вологи (ДАВ).
Вологість в'янення залежить від ґрунтових умов, головним чином від механічного складу ґрунту. На супісках вона становить 1‑3%, на суглинках – 3‑10, на глинистих ґрунтах –10‑15%. Вона близька до полуторної максимальної гігроскопічності ґрунту. Часто вологість в'янення залежить від властивостей рослини, її віку.
У міру росту і розвитку рослини коренева система її також збільшується в розмірах, заглиблюється і охоплює все більший обсяг ґрунту Коріння рослин досягають різної глибини. Наприклад, люцерни і буряків проникає на глибину 3‑5 м, пшениці й кукурудзи – до 2, проса і гороху – близько 1 м. Неглибоко заходять у ґрунт корені капусти, цибулі. Проте основна маса коріння зосереджена на значно меншій глибині ґрунту: у однорічних рослин – у верхній половині кореневмісною шару, у багаторічних трав – у верхній третині. Цей шар ґрунту, в якому розміщено до 90% кореневих волосків і найтонших коренів, найбагатший поживними речовинами і заселений мікроорганізмами, називається активним. І Його можна розглядати також як шар активного водоспоживання.
Глибина активного шару змінюється залежно від фаз розвитку рослин. При проведенні вегетаційних поливів вологість активного шару підтримують на заданому рівні. Кореневмісний шар або деяка його частина, розміщені глибше активного шару ґрунту, зволожуються в разі потреби допосівними поливами.
Надходження до рослини ґрунтової вологи і розчинених у ній поживних речовин – складний біологічний процес. Він у значній мірі визначається величиною всмоктувальної сили клітин кореневих волосків і осмотичним тиском клітинного соку.
Всмоктувальну силу клітин (S) можна визначити за формулою:
S = P – T,
де Р ‑ осмотичний тиск клітинного соку; Т – тургорний тиск клітин.
Осмотичний тиск і всмоктувальна сила клітин різні у різних видів рослин. Вони змінюються також з віком рослин і залежно від зовнішніх умов. В міру старіння рослинного організму осмотичний тиск звичайно підвищується. Чим вищі запаси води в ґрунті і більше її міститься в тканинах рослин, тим нижчий осмотичний тиск і всмоктувальна сила.
При зменшенні водного запасу ґрунту всмоктувальна сила листків і коріння підвищується, але при цьому збільшується і всмоктувальна (водоутримна) сила ґрунту. Внаслідок цього засвоєння води корінням обмежується. Найбільш інтенсивно вона поглинається коренями при оптимальній вологості ґрунту, сприятливій для швидкого росту рослин.
Про швидкість засвоєння води кореневою системою можна робити висновок за кількістю пасоки, що подається коренями в надземні органи рослин.
При поливі поглинання води корінням збільшується. В міру старіння рослин швидкість надходження води в клітини кореня зменшується незалежно від умов водопостачання.
Надходження води в рослини залежить також від припливу кисню до рослинних клітин, оскільки з підвищенням інтенсивності дихання посилюється надходження води в кореневу систему рослин. У дослідах Г М. Гриньової (1963) кукурудза і соняшник поглинали воду в анаеробних умовах у декілька разів повільніше, ніж при нормальній аерації. Таким чином, надмірне зрошення, яке створює анаеробні умови, несприятливе для надходження води в рослини.
Д.А. Сабінін, А.М. Алексєєв (1969) та інші встановили тісний зв'язок між поглинанням води і живленням рослин. При оптимальній кількості поживних речовин відмічено найбільшу швидкість поглинання води, при недостатньому живленні вона зменшується. Особливо затримується цей процес при азотному голодуванні.
Корисний, або легкодоступний, запас ґрунтової вологи рослина споживає згідно з фазами розвитку, органотворними процесами, формуванням і діяльністю кореневої системи.
У посушливих умовах рослини в міру розвитку використовують вологу із більш глибоких шарів ґрунту. В зв'язку з цим при недостатній кількості опадів спостерігається висушування спочатку орного, потім підорного і пізніше більш глибоких шарів кореневмісного шару ґрунту .
У початкові фази кількість вологи, яка споживається рослинами, невелика внаслідок слабкого розвитку випаровуючої (листкової) поверхні і відносно невеликої сухості повітря на початку вегетаційного періоду. Між тим інтенсивність транспірації, тобто випаровування води з одиниці площі листка за одиницю часу, може бути високою, оскільки поряд з продиховим дуже розвинуте кутикулярне випаровування.
У подальшому в міру збільшення площі листків, підвищення температури і сухості повітря, а також посилення суховіїв кількість води, що споживається з ґрунту і витрачається рослинами, збільшується. При цьому кутикулярне випаровування знижується, оскільки зменшується проникність кутикулярного шару для води.
Період найбільшого споживання ґрунтової вологи рослинами нерідко припадає на фази цвітіння і початок розвитку зав'язей. До кінця вегетації воно зменшується. Запас ґрунтової вологи зменшується також внаслідок випаровування з поверхні ґрунту.
Витрата ґрунтової вологи через транспірацію і ґрунтове випаровування становить сумарне випаровування (синоніми – евапотранспірація, сумарне водоспоживання). Для сільськогосподарського виробництва цей показник має більш важливе значення, ніж визначення величини транспірації, оскільки він відображає дійсне становище, в якому створюється водний режим ґрунту і рослин. Джерела сумарного випаровування – природні запаси води в ґрунті, опади, зрошувальна вода, підґрунтові води (при близькому їх стоянні).
Сумарне випаровування, або сумарна витрата води полем, складається з таких величин:
![]()
де åі – сумарне випаровування, м3/га; Wз – запас вологи в кореневмісному шарі під час сівби, м3/га; Wу ‑ запас вологи кореневмісного шару на час збирання, м3/га; Ок – корисна частина (що. проникає в ґрунт) опадів, м3/га; Мз – зрошувальна норма, м3/га; К – кількість води капілярної зони, що використовується при незначному рівні підгрунтових вод, м3/га.
Сумарне випаровування за вегетацію даної культури, що припадає на одиницю одержаного врожаю, називається коефіцієнтом водоспоживання (синоніми ‑ коефіцієнт сумарного випаровування, коефіцієнт евапотранспірації). Він показує, скільки води витрачено ґрунтом і рослиною на утворення валової одиниці врожаю:
![]()
де KB ‑ коефіцієнт водоспоживання, м3/т; åі ‑ сумарне випаровування, м3/га; У – урожай, ц/га.
Вплив зрошення на величину і якість урожаю. Поліпшуючи водоспоживання рослин і змінюючи ґрунтові й мікрокліматичні умови, зрошувальна вода суттєво впливає на ріст і розвиток, а також на продуктивність сільськогосподарських культур. Вільне надходження води і вуглекислого газу до рослин підвищує асиміляцію в 5‑8 разів, а іноді і більше.
Комплекс сприятливих для рослин умов, які створюються на зрошуваних землях, сприяє нормальному формуванню кореневої системи, доброму росту й розвитку рослин. У результаті врожайність при зрошенні підвищується в 3‑4 рази і більше.
Зрошення впливає не тільки на величину врожаю, але і на його якість. Цукристість коренеплодів цукрових буряків звичайно зменшується з підвищенням вологості ґрунту (табл. 6).
6. Вплив режимів зрошення на врожайність цукрових буряків, цукристість
коренеплодів і збір цукру (Вороній Н. Г., 1989)
|
Поливи при вологості ґрунту, % НВ |
Урожайність, ц/га |
Вміст цукру в корінні, % |
Збір цукру, ц/га |
|
Без зрошення |
102 |
22,3 |
22,8 |
|
60 |
302 |
19,5 |
58,9 |
|
70 |
547 |
18,8 |
102,9 |
|
80 |
560 |
18,4 |
102,9 |
Цукристість коренеплодів знижують головним чином пізні поливи при температурі нижче 20°С, тому за 3‑4 тижні до збирання буряків поливи припиняють. Проте збір цукру з 1 га при зрошенні за рахунок росту врожайності різко збільшується. Цукристість при зрошенні піддається регулюванню – її підвищують фосфорні и калійні добрива. Позитивна сторона зрошення – зменшення в коренеплодах вмісту «шкідливого азоту», що знижує вихід кристалізаційного цукру.
Вміст крохмалю в бульбах картоплі при підвищенні зрошувальної норми може знижуватися. Проте врожайність бульб у цьому
випадку настільки збільшується, що валові збори крохмалю при зрошенні виявляються все-таки набагато вищі, ніж без нього. Підвищують вміст крохмалю фосфорні добрива.
Вміст клітковини, особливо в стеблах зернових культур, нерідко збільшується з підвищенням зрошувальної норми. При цьому може дещо зменшуватися відношення зерна до соломи. Воно, однак, піддається регулюванню шляхом встановлення відповідних норм висіву, добрив, строків сівби тощо.
Щодо вмісту жиру, то кількість його при поливах помітно підвищується в соняшнику, сої, рицині і знижується в ефіроолійних культурах.
Багато авторів відзначають, що при зрошенні знижується вміст протеїну в насінні пшениці, олійних рослин та інших культур. Причиною цього найчастіше є неправильне поєднання режиму зрошення і мінерального живлення. Ці ж причини можуть викликати підвищення кислотності помідорів.
Роль добрив у підвищенні якості зерна поливної озимої пшениці наведено в таблиці 7.
7. Вплив зрошення і удобрення на врожай і якість зерна
озимої пшениці (Кириченко В.П.)
|
Показники обліку і одиниці виміру |
Без зрошення і добрив |
Зрошення | |
|
без добрив |
на фоні добрив | ||
|
Урожайність зерна, ц/га |
17,3 |
33,4 |
50,4 |
|
Маса 1000 зерен, г |
40,2 |
42,0 |
45,0 |
|
Натура зерна, г/л |
790 |
805 |
809 |
|
Білок, % |
14,51 |
12,42 |
14,40 |
|
Сира клейковина, % |
34,1 |
23,7 |
30,7 |
При сумісному застосуванні зрошення і добрив за якістю продукції озима пшениця сорту Безоста 1 входить до розряду сильних.
Вміст білка і клейковини в озимої пшениці в значній мірі підвищують позакореневі підживлення азотом. Його можна вносити разом з поливною водою у фази виходу в трубку і колосіння. Підживлення у фазі колосіння сорту Одеська напівкарликова в дослідах Інституту зрошуваного землеробства УААН дало можливість підвищити вміст білка в зерні до 14,3%, клейковини – до 40%, об'єм хліба – 915 см3 з відмінною його оцінкою.
Підживлення азотом з поливною водою підвищувало вміст білка в зерні озимого ячменю від 8,89 до 12,25%, азот разом з фосфором (N180P120) У зерні кукурудзи – від 9,30 до 10,75% і (N60P60) в зерні сої – від 31,6 до 32,6%.
Безперечно, якість урожаю на поливних землях залежить також від сорту або гібриду. Окремі сорти м'якої і твердої пшениці, які мають високу «силу» борошна та інші позитивні якості, зберігають і поліпшують їх при зрошенні, якщо створений відповідний агротехнічний фон. Добрива в значній мірі поліпшують хлібопекарські якості борошна озимої пшениці.
Правильне застосування добрив та інших заходів сумісно з оптимальним режимом зрошення поліпшує хімічний склад, розміри і масу зерна пшениці, смакові якості овочевих культур.