4.1. Теоретичні основи селекції
Розведення — це наука про створення і розмноження сільськогосподарських тварин і птиці та поліпшення їхніх спадкових якостей. Воно передбачає розроблення теоретичних основ та практичних прийомів племінної роботи у тваринництві. Під племінною роботою у вівчарстві розуміють систему організаційно- зоотехнічних заходів щодо поліпшення породних і продуктивних якостей овець.
Виробничий процес у вівчарстві має трикомпонентну структуру: продукція, селекція і технологія. Селекція забезпечує формування спадкових задатків бажаного комплексу ознак продуктивності і адаптації овець. Вона супроводжується зміною поколінь організмів на основі оцінки індивідуальних показників продуктивності і здоров’я тварин, що формуються у фотогенезі за часовими періодами процесу виробництва. Результатами селекції визначаються якісні, кількісні та економічні показники у формі собівартості та реалізаційної ціни одиниці продукції. Продукція і селекція мають широкий спектр взаємозалежних зв’язків. Вимоги ринку до якісних характеристик продукції, а вони на фоні існуючого попиту формують реалізаційну ціну одиниці продукції, становлять основу нормативної бази селекції за якісними показниками продуктивності овець.
Селекція забезпечує поліпшення спадкових задатків продуктивності та здоров’я овець, її теоретичні основи є практично однаковими для всіх видів сільськогосподарських тварин і ґрунтуються на закономірностях мінливості в популяції. Особливості методів племінної роботи у вівчарстві зумовлені специфікою продукції овець — вовни, смушків, овчин. Успішне ведення її потребує від спеціалістів знань основ генетики, практичного володіння прийомами оцінювання, добору і підбору тварин, вмінням кваліфіковано аналізувати і планувати розвиток галузі. В цілому селекція, як одна з трьох складових вівчарства включає об’єкти роботи і методи. Об’єкти — це вівці з часу одомашнення до сучасних виробничих напрямів вівчарства, а методи — практична реалізація закономірностей селекції. Селекція — це складний комплекс зоотехнічних заходів, спрямованих на якісне перетворення і раціональне використання існуючих, а також створених нових порід, типів, ліній та їх кросів. Як визначав М.І. Вавилов, селекція — це еволюція живих організмів, яка спрямовується волею людини.
Установлено, що всі рослини і тварини складаються із клітин. Кожна клітина має складну біотехнічну структуру і складається із клітинної оболонки, цитоплазми та ядра. Ядроклітини в період спокою має сітчасту структуру із тонких ниточок. Під час поділу клітини ниточки скорочуються і набирають форми паличок, здатних фарбуватися, їх видно під мікроскопом, і називаються вони хромосомами. Головне призначення хромосом — бути носіями генів. Гени — матеріальні часточки, що лежать в основі життєдіяльності організму і є відповідальними за формування окремих його ознак.
Кожен вид тварин характеризується своїм відповідним набором хромосом. В овець, наприклад, їх 54. При цьому в кожному наборі хромосом можна знайти по дві подібні. Відмінності за кількістю хромосом між баранами і вівцематками немає. Незважаючи на низький рівень вивчення каріотипу, відомо багато факторів спадкових порушень кількісного або морфологічного складу хромосом у тварин. Такі відхилення можуть бути причиною зміни деяких продуктивних ознак і прояву спадкових аномалій, що завдають значних збитків виробництву.
Установлено, що ріст і розвиток живого організму відбувається на основі поділу та утворення нових соматичних і статевих клітин. У процесі розмноження хромосоми соматичних клітин поділяються і розходяться в однакових кількостях у дочірні клітини. Розмноження клітин за таким принципом називається мітозом. Тип розмноження статевих клітин називається мейозом. На відміну від мітозу, він завершується утворенням двох дочірніх клітин із однієї материнської з половинним набором хромосом. У результаті цього всі яйцеклітини ярок мають по одній Х-хромосомі, а спермії баранів можуть бути з Х або У-хромосомою. При поєднанні батьківської і материнської Х-хромосом розвиватиметься організм ярочки, а при поєднанні яйцеклітин з Х-хромосомою та спермія з У-хромосомою — потомство чоловічої статі. Отже, відмінність між сперміями, половина з яких має X, а решта — У-хро- мосому, є біологічним механізмом, який зрівноважує подібне народження ягнят різної статі.
Науковими дослідженнями встановлено, що показники розвитку, багатоплідності, м’ясної і молочної продуктивності визначаються багатьма факторами, які можна поділити на дві категорії: середовищні і спадкові. До факторів середовища, які істотно впливають на продуктивність тварин, належить якість кормів, повноцінність раціонів і рівень годівлі, мікроклімат приміщення та кліматичні особливості зони, технологія утримання тварин, кваліфікація обслуговуючого персоналу, організація виробничих процесів. Шляхом раціонального використання зазначених факторів можна порівняно швидко і значною мірою підвищити продуктивність тварин, але не вище рівня, зумовленого спадковими можливостями їхнього організму.
Природою так установлено, що кожна жива істота залишає подібне собі потомство. Потомству від батьків передаються і характерні породні особливості. Явище давати подібне собі потомство називають спадковістю.
Проте як би споріднені тварини не були схожі між собою, кожна з них має свої особливості. Це явище у споріднених тварин називають мінливістю.
Частка мінливості, зумовлена генетичними факторами, називається успадкуванням і позначається Я2. Коефіцієнт успадкування визначається різними методами:
♦ подвоєнням коефіцієнта кореляції г між батьками і нащадками —![]() (батьки — нащадки);
(батьки — нащадки);
♦ подвоєнням коефіцієнта регресії![]() між батьками і нащадками —
між батьками і нащадками —![]() (батьки — нащадки);
(батьки — нащадки);
♦ однофакторним дисперсійним комплексом та ін.
Коефіцієнти успадкування можуть змінюватися від 1 до 100 % або від 0 до 1 залежно від методу їх розрахунку (табл. 4.1).
|
Таблиця 4.1. Відносна оцінка успадкування в овець (за К. Террілом, 1968)
|
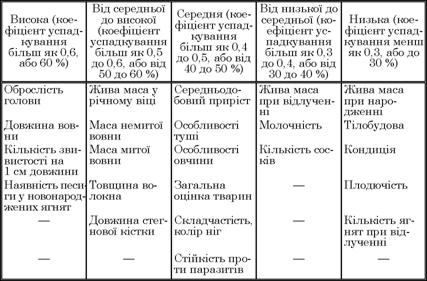
Чим вищий показник цього коефіцієнта, тим більші можливості передачі ознак від батьків нащадкам. За даними табл. 4.1, господарсько-корисні ознаки овець успадковуються по-різному. Порівняно низькими коефіцієнтами успадкування характеризуються репродуктивні якості тварин (Я2 до 30 %). Це свідчить про те, що селекція батьків на підвищення, наприклад, плодючості у нащадків за цими ознаками є явно малоефективною, тобто репродуктивні якості овець треба підвищувати, поліпшуючи умови їх годівлі та утримання.
Рівень показників успадкування відгодівельних якостей, які характеризують ріст тварин, середній (Я2 у межах 40 — 50 %). В умовах виробництва середовище впливає на виявлення цих ознак більше, ніж генетичні фактори, але передача їх нащадкам майже в 1,5 — 2 рази більша, ніж ознак, які характеризують репродуктивні якості овець.
Ще більшою є можливість селекції овець за настригом вовни та її довжиною. Коефіцієнти успадкування цих ознак часто коливаються в межах 50 — 60 % і більше. Відповідно вовнову продуктивність ефективніше змінювати за рахунок не повноцінності і рівня годівлі, а генетичних факторів.
В овець планових порід України вивчено успадкування більш як 20 ознак (табл. 4.2).
|
Таблиця 4.2. Показники успадкування найважливіших господарських ознак овець
|
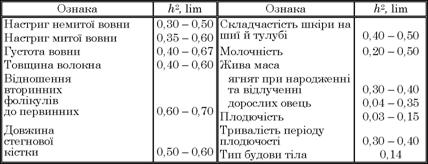
Коефіцієнт успадкування ознак у практиці племінної роботи враховують для певної популяції або стада овець. Його не можна механічно переносити на інше стадо, бо кожна популяція мас свої особливості. Високий показник успадкування тієї чи іншої ознаки завжди буває у стаді з більшою різноманітністю генотипової інформації за ознаками, що вивчаються. Аналіз цих даних дає змогу визначити загальну закономірність в успадкуванні ознак. Ті ознаки тварин, розвиток яких значною мірою залежить від умов життя, мають нижчий коефіцієнт успадкування. До таких ознак належать насамперед вовнова, молочна і м’ясна продуктивність. Якісні ознаки тварин, на яких менше впливають фактори зовнішнього середовища (наприклад, довжина й густота вовни, складчастість шкіри, відношення вторинних фолікулів до первинних та ін.), мають більш високий показник успадкування, що, в свою чергу, свідчить про те, що в їх загальній мінливості частка спадкової різноманітності вища, ніж в ознак з низькою спадковістю.
Кожна ознака організму виявляється в умовах, сприятливих для його розвитку. Якщо зовнішні умови несприятливі для нормальної життєдіяльності тварин, то генотипові якості не виявляються. Показник успадкування буди нижчим порівняно з його величиною за оптимальних екологічних умов. М.Ф. Іванов з великої кількості факторів зовнішнього середовища виділяв годівлю та умови утримання тварин. Він вважав, що спадкові якості виявляються повною мірою тільки за найсприятливіших умов годівлі, утримання й догляду. Виходячи з цього, він настійно рекомендував утримувати відтворне стадо овець, особливо молодняк, за оптимальних умов зовнішнього середовища.
Слід пам’ятати, що зі збільшенням генетичної однорідності особин конкретного стада коефіцієнт успадкування зменшується і умови середовища стають основними факторами, що зумовлюють відмінності між тваринами за продуктивними якостями. У разі посилення однорідності середовища, а в деяких випадках і поліпшення умов, особливо умов годівлі, коефіцієнт успадкування збільшується за рахунок зростання значущості індивідуальної відмінності між тваринами, зумовленої спадковістю.
Через різне співвідношення спадкових і неспадкових факторів, які визначають мінливість господарсько-корисних ознак, їх успадкування в різних стадах неоднакове. При цьому для вибору того чи іншого методу добору і прогнозування його ефективності коефіцієнти успадкування треба визначати для тварин конкретного стада.
Безумовно, в кожному конкретному випадку можливі незначні відхилення від загальних закономірностей успадкування ознак, але загалом вони виявляються чітко. Знаючи, як успадковується та чи інша ознака, можна відбирати для наступного розмноження тільки більш продуктивних тварин і передбачати якість потомства від різних варіантів парування, а отже, раніше планувати їх.
Генетично розрізняють два види ознак у тварин: якісні й кількісні. До якісних ознак в овець належать колір вовни, ніг, голови, форма хвоста та інші анатомо-морфологічні особливості організму. Визначаються вони одним або порівняно невеликою кількістю генів. Селекція за цими ознаками досить проста, і її результати можна передбачити з високою точністю. Кількісні ознаки визначають господарсько-корисні якості тварин і успадковуються полі- генно. До них належить сукупність анатомічних, фізіологічних, біохімічних особливостей організму. Кожна із них, у свою чергу, детермінована багатьма генами, генними системами та їхньою взаємодією залежно від умов середовища. Полігенність кількісних ознак зумовлює різноманітність форм їх успадкування, яке відбувається за адаптивним або проміжним типом, а це дуже ускладнює прогнозування бажаних результатів. Селекція тварин за кількісними ознаками набагато складніша від якісної і потребує урахування багатьох факторів, що пов’язано з великими затратами праці й часу.
Організм розвивається як єдине ціле під впливом спадковості та умов середовища. Тому всі функції організму взаємопов’язані між собою. Зміна функцій одних органів і тканин веде до зміни функцій інших органів і тканин. Таке явище називається кореляційною мінливістю. Кореляція (взаємозв’язок) між господарсько-корисними ознаками буває фенотиповою і генетичною. Фено- типова кореляція показує ступінь відносної мінливості феноти- пів, а генетична — як змінюється одна ознака у нащадків, якщо проводити відбір батьків з другою ознакою, що взаємопов’язана з першою. Виражається кореляція величиною від 0 до 1. При кореляції, що дорівнює 0, коефіцієнт кореляції г нульовий, а при кореляції, що дорівнює 1, називається функціональним (зі зміною однієї ознаки інші змінюються пропорційно). Коефіцієнт кореляції до 0,4 вважається низьким, від 0,4 до 0,7 — середнім і вище 0,7 — високим.
Взаємозв’язок між ознаками буває позитивним (збільшення або зменшення однієї ознаки супроводжується такою самою зміною іншої) і негативним (збільшення однієї ознаки спричинює зменшення іншої і навпаки).
За високої генетичної кореляції між двома ознаками відбір тварин за однією з них приведе до генетичної зміни обох ознак. Прикладом може бути висока від’ємна кореляція (г = —0,73) між швидкістю росту та ефективністю використання корму вівцями. Багато наукових даних свідчать про те, що відбір овець протягом кількох поколінь на збільшення росту одночасно сприяє зменшенню витрат корму на 1 кг приросту. Це означає, що як фізіологічно, так і генетично ці дві ознаки корелюють між собою і для їх поліпшення достатньо вести селекцію за однією з них. Якщо між ознаками немає кореляції (вона близька до нуля), то для їх генетичного поліпшення проводиться незалежний відбір.
За наявності небажаної від’ємної кореляції між ознаками, наприклад, між довжиною і густотою вовни, відбір проводиться з урахуванням обох ознак, оскільки незалежний відбір за однією з них призводить до генетичного погіршення іншої.
|
Таблиця 4.3. Генетичні кореляції між основними господарсько-корисними ознаками овець
|

Як видно з табл. 4.3, в овець на відгодівлі збільшення енергії росту зумовлює зниження витрат корму на одиницю приросту маси тіла та розвиток вовни.
Доведено, що генетичні параметри стада характеризуються також повторюваністю, що вказує на стабільність ознак у постембріональний період розвитку тварин. Використовуючи коефіцієнти повторюваності, вже за результатами ранньої оцінки тварин можна прогнозувати їхню наступну продуктивність. Тобто коефіцієнти повторюваності, як і коефіцієнти успадкування, можуть бути основою для прогнозування ефективності добору.
Підвищення селекційної ознаки у нащадків порівняно з вихідними батьківськими формами прийнято називати генетичним зрушенням. Останні залежать від селекційного тиску, селекційного диференціала, коефіцієнта спадковості та швидкості зміни поколінь. Під селекційним тиском розуміють відсоток вибраку- ваних тварин після проведеного оцінювання їх. Чим менше відбирають тварин для подальшого відтворення, тим вищий селекційний тиск. Якщо, наприклад, оцінили 500, а відібрали 250 голів, то селекційний тиск становитиме 50 %, а якщо відібрали 100 голів, то 80 % і т.д. Перевищення селекційною ознакою у тварин, залишених для наступного розмноження, усіх оцінених по стаду ознак називаютьселекційним диференціалом, який обчислюють за формулою
![]()
де![]() — середній показник ознаки по всіх відібраних тваринах;
— середній показник ознаки по всіх відібраних тваринах; ![]() — середній показник по всіх тваринах без впливу відбору.
— середній показник по всіх тваринах без впливу відбору.
Приклад. На племінній фермі, де утримуються вівці породи прекос, є 2400 маток, від яких щороку вирощують 2400 голів молодняку, в тому числі 1200 ярок і 1200 баранців. Настриг вовни від однієї матки становить 4,2 кг, а
від однієї ярки — 3,2 кг. Перспективним планом роботи зі стадом передбачено посилити селекцію щодо вовнової продуктивності, а тому щорічне вибраковування вівцематок проводиться не за віком, а за настригом вовни і становить 20 %. Баранів вибраковують 17 %. Відповідно до планового завдання щороку вибраковують 480 маток з низьким настригом вовни. В стаді залишається 1920 маток з кращим настригом (наприклад, середній настриг становить 4,4 кг, або на 0,2 кг більший, ніж у маток усього стада).
Для обчислення селекційного диференціала матерів наведена вище формула матиме такий вигляд:
![]()
де![]() — селекційний диференціал по матерях;
— селекційний диференціал по матерях;![]() — селекційний настриг від відібраних на плем’я маток;
— селекційний настриг від відібраних на плем’я маток;![]() — середній настриг від усіх маток стада.
— середній настриг від усіх маток стада.
Підставивши в цю формулу наведені у прикладі дані про настриг вовни, матимемо
![]()
Для ремонту вибракуваних маток відбирали 480 ярок із середнім настригом від однієї 3,5 кг. їхній настриг перевищував середній показник по всіх ярках на 0,3 кг. Отже, загальна вовнова продуктивність маточного поголів’я без впливу відбору становитиме
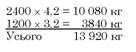
Поділивши загальну кількість вовни на кількість маток і ярок, які є в господарстві, матимемо середній настриг по маточному стаду. В нашому прикладі він становить 13 920 : 3600 = 3,87 кг. Після цього обчислюємо загальну кількість вовни, одержану від маток і ярок, відібраних для відтворення:
![]()
Поділивши суму 10 128 на поголів’я відібраних тварин (2400), одержимо середню величину 4,22 кг.
Отже, у нашому прикладі при коефіцієнті відбору ярок 40 % і маток 20 % селекційний диференціал по маточному стаду ферми становитиме
![]()
Вихідні дані для визначення селекційного диференціала баранів- плідників такі: для осіменіння 2400 маток з урахуванням розведення по лініях на фермі має бути 24 основних плідники. Для їх ремонту щороку потрібно 4, а разом із пробниками — 24 молодих баранці. Середній настриг вовни основних баранів у річному віці — 5,5 кг, а решти для подальшого відтворення — 6 кг, тобто на 0,5 кг більший порівняно з усіма баранами.
Від вирощуваних на фермі 1200 баранців на одну голову одержано в середньому 4,1 кг вовни, а по групі відібраних для ремонту основного стада 24 плідників — 6,2 кг. Різниця на користь відібраних — 2,1 кг. Загальна вовнова продуктивність усіх баранів стада складатиметься із настригу від баранів основного стада і молодняку. Розрахунок здійснюють так:
|
|
Поділивши отриману величину на кількість баранів (1224), одержимо середній настриг вовни від одного барана. Він становитиме 5052 : 1224 = = 4,13 кг. Тепер визначимо настриг вовни від усіх залишених на плем’я баранів:
|
|
Середня продуктивність залишених для відтворення баранів буде такою:
|
|
Селекційний диференціал для плідників визначимо за формулою
|
|
|
|
|
|
Загальний селекційний диференціал відтворення поголів’я стада (матки, ярки та барани) визначимо за формулою
|
|
де![]() — загальний селекційний потенціал по всьому стаду;
— загальний селекційний потенціал по всьому стаду;![]() — селекційний диференціал по баранах;
— селекційний диференціал по баранах;![]() — селекційний диференціал по маточній групі.
— селекційний диференціал по маточній групі.
У нашому прикладі він становитиме
|
|
Отже, загальний селекційний диференціал стада овець складається із півсуми показників селекції: диференціала маточної групи стада і диференціала баранів усього стада ферми.
Відповідь на селекцію. Очікувану відповідь на селекцію за відповідними ознаками, що характеризують продуктивність тварин, визначають за формулою
![]()
де![]() — очікувана відповідь на селекцію;
— очікувана відповідь на селекцію;![]() — коефіцієнт успадкування;
— коефіцієнт успадкування;
![]() — загальний селекційний диференціал стада ферми.
— загальний селекційний диференціал стада ферми.
Для одержання відповіді на селекцію по нашій фермі на 2400 маток нам бракує показника успадкування по настригу вовни. Припустімо, що коефіцієнт успадкування по матках та їхніх нащадках становить 0,46. Тоді очікувана відповідь на селекцію не одне покоління
![]()
Оскільки на фермі прийнято замінювати маточне поголів’я через 5, а баранів через 6 років, така зміна поколінь у нашому стаді відбудеться за 5,5 року. З урахуванням цього визначимо очікувану відповідь на селекцію за один рік. Для цього показник, що виражає селекцію на одне покоління, поділимо на кількість років, протягом яких відбувається зміна поколінь:
0,418 : 5,5 = 0,076 кг.
Отже, у нашому стаді вовнова продуктивність щороку підвищуватиметься на 0,076 кг, а за одне покоління — на 0,417 кг. Через одне покоління середній настриг по маточному стаду ферми становитиме 4,2 + 0,418 = 4,618 кг.
Зазначимо, що в багатьох господарствах показникам селекційного тиску і достатнього селекційного диференціала приділяється недостатньо уваги. Часто на племінних фермах ірепродукторах вирощують таку кількість ремонтного молодняку, якої вистачає тільки на оновлення основного стада. При вирощуванні недостатньої кількості овець після оцінки селекційний диференціал мало відрізнятиметься від нульового значення. У такому разі нічого очікувати генетичного зрушення ознаки в наступному поколінні. Якісні показники стада в кращому разі залишаться на попередньому рівні.
На ефективність селекційного процесу впливає і таке явище, як інтервал між поколіннями. Він прискорює або уповільнює селекційний процес і є основною одиницею часу в селекції. Швидка зміна поколінь за інших однакових умов є основним фактором прискорення селекційного процесу. Інтервал між поколіннями дорівнює періоду від народження самої тварини до народження її нащадків, залишених для розведення.
Величина генераційного інтервалу залежить переважно від системи розведення, методів оцінки племінної цінності тварин і технології тваринництва. Відбір тварин з високим і вірогідним рівнем племінної цінності підвищує селекційний диференціал,
але водночас збільшує їхній генераційний інтервал. Так, якщо ярок уперше парували у 18-місячному віці і відбирали на плем’я потомство при народженні, то часовий проміжок між поколіннями становитиме 23 міс. У практиці багатьох господарств він досягає 4,0 - 4,5 року.
Оцінюючи генетичні основи селекції, слід враховувати форми прояву спадкових якостей у потомстві — проміжне успадковування, гетерозис та інбредну депресію.
Проміжне успадковування господарсько-корисних ознак спостерігається у племінних і товарних стадах за неспорідненого внутрішньопородного розведення. Якщо врахувати, що кількість ознак характеризується адаптивним успадковуванням, то за типового для багатьох господарств невисокого рівня селекції середня продуктивність стада здебільшого зберігається. За доброї організації вирощування ремонтного молодняку, цілеспрямованого добору і підбору тварин, як правило, забезпечується підвищення продуктивності якостей стада у межах спадкових можливостей породи або типу.
Гетерозисом у тваринництві називають явище переважання нащадків першого покоління над батьківськими формами за окремими господарськими ознаками, що виникає в результаті застосування відповідних методів схрещування. У вівчарстві гетерозис звичайно спостерігається в різних варіантах схрещування і виявляється у високій життєздатності та витривалості нащадків, а часто й у вищій продуктивності їх порівняно з батьківськими формами при розведенні «в собі». Залежно від особливостей і по- єднаності порід, яких використовують для схрещування, а також від типів і ліній гетерозис може виявлятися у збільшенні багатоплідності, великоплідності ягнят, їх збереженні і швидкостиглос- ті, поліпшенні вовнової, м’ясної і смушкової продуктивності, у зменшенні витрат кормів на одержання продукції.
Установлено, що проміжне успадкування може спостерігатися не тільки при внутрішньопородному розведенні, а й при схрещуванні, а гетерозис, у свою чергу, — і при паруванні чистопородних баранів та вівцематок. За допомогою аналізу даних схрещування виявлено таку тенденцію: за частотою і рівнем прояву гетерозису провідними виявляються ознаки, які характеризують відтворну здатність овець, потім — вовнові і далі м’ясні.
Отже, чим вищий коефіцієнт успадковування, тим меншою стає гарантія одержання гетерозису за цією ознакою при проведенні внутрішньопородних або міжпородних парувань.
Інбредну депресію можна вважати протилежною формою гетерозису. Вона є результатом близьких споріднених парувань, особливо при багаторазовому і безсистемному їх застосуванні. У вівчарстві інбредна депресія може виявлятися в ослабленні конституції, зниженні плодючості, появі каліцтв та пригніченні росту і розвитку тварин.
На підставі аналізу багатьох даних про інбридинг-депресію у тваринництві можна зробити висновок, що сила прояву депресії залежить від виду, породи і лінійної належності тварин. Існує правило, за яким ступінь прояву інбредної депресії прямо пов’язаний із плодючістю тварин. Для малоплідних тварин він більший, ніж за тісного інбридингу, і є складним селекційним методом, який потребує високої кваліфікації виконавців. Він ґрунтується на чіткому доборі і жорсткому вибракуванні небажаних тварин та комбінацій їх одержання. Безсистемність у застосуванні тісного інбридингу може спричинити негативні наслідки.
Однак історія зоотехнії поступово накопичувала дані, що отримані з використанням інбридингу тварини не тільки не відстають від аутбредних тварин, а навіть переважають їх. Відомо, що Беквел створював нові породи на основі інбридингу будь-яких ступенів з метою закріплення у нащадків високоцінних якостей родоначальників. Таким методом створено лейстерську породу овець.
Пізніше було встановлено, що успіх при використанні інбридингу визначається певними обставинами, головні з яких такі:
♦ цінність родоначальника, на якого проводиться інбридинг. Якщо родоначальник не вирізняється високою племінною цінністю, то будь-який інбридинг на нього буде негативним фактором. Інбридинг на посереднього загального нащадка призводить до погіршення результатів, оскільки він не компенсований цінною спадковістю, інбредна депресія при цьому виявляється досить виразно;
♦ ступінь інбридингу. Вважають, що тісний інбридинг доцільно використовувати на перших етапах створення нової породи, коли кількість відібраних тварин, що відповідають цільовому стандарту, незначна, і потрібно закріпити цінні якості родоначальника.
Тісний інбридинг при розведенні за лініями використовується рідко, переважно тільки у роботі з унікальними тваринами.
У племінній роботі є ще такі поняття, як генотип, фенотип, домінування, рецесивність, гомозиготність, гетерозиготність тощо.
Під генотипом розуміють сукупність усіх генів, які зумовлюють спадковість тварин, тобто контролюють ріст, розвиток і функціонування організму від ембріона, народження тварини до її вибракування або природної смерті.
Фенотип — це сукупність усіх ознак і властивостей організму, які доступні для спостереження та аналізу або утворилися внаслідок взаємодії генотипу з умовами навколишнього середовища.
Середовище — комплекс негенетичних факторів (рівень годівлі, умови утримання, мікроклімат у приміщенні, фізіологічний стан, вік тварин та ін.), які впливають на розвиток і функціонування організму. У різних умовах середовища по-різному реалізується генотип у вигляді фенотипових властивостей тварин — рівень вовнової, м’ясної, молочної продуктивності та ін.
Під домінуванням розуміють переважання в прояві однієї ознаки (гена) над іншими.
Рецесивний стан ознаки (гена) показує, що ця ознака пригнічена, перебуває у прихованому стані.
Гомозиготним вважають генетично однорідний організм, що утворився в результаті злиття гамет з подібними за даною ознакою генами і тому продукує спадково подібні клітини.
Гетерозиготним вважається генетично неоднорідний організм, що утворився в результаті злиття гамет з різними генами і тому продукує спадково неподібні клітини.
Отже, успіх застосування сучасних досягнень науки і практики в селекції залежить від безпосередньої роботи спеціаліста. Тільки цілеспрямоване вирощування молодняку, добір і підбір батьківських пар з наступною оцінкою результатів їх парування та повторення правильних поєднань можуть забезпечити стійкі результати продуктивності тварин і вдосконалення всього стада в бажаному напрямі.