1.4.1. Походження та еволюція свійських овець
За зоологічною класифікацією, вівці належать до класу ссавців, підкласу плацентарних, загону парнокопитних, підзагону жуйних, родини порожнисторогих, підродини вівцекози, роду вівці, виду дикі вівці, підвиду свійські вівці. Підродина вівцекози має п’ять родів (кози, тари, нахури, гривасті барани, вівці). Дикі вівці мають два види — гірські барани і товстороги. Гірські барани поділяються на муфлоноподібні та аргалоподібні підвиди, товстороги — на азіатські й північноамериканські.
Вівця є однією з перших тварин, яку людина приручила й одомашнила. У навколишній природі було багато диких видів овець, які і тепер поширені в Європі, Азії та Північній Америці. На основі археологічних, антропологічних, зоологічних та морфологічних досліджень, порівняльного вивчення багатьох особливостей диких і свійських видів багато дослідників (К. Келлер, А. Нерінг, Л. Адамець, П.М. Кулешов, Є.А. Богданов, М.Ф. Іванов, С.М. Боголюбський, В.І. Цалкін та ін.) дійшли висновку, що одомашнення овець здійснювалося в різних районах планети — Південній Європі, Передній Азії, Північній Африці, Малій Азії, Середній і Центральній Азії. Існує думка, що людина одомашнила 3 види, або раси, диких баранів — муфлонів, аркарів, або урі- алів, архарів, або аргалів, тобто стверджується поліцентризм одомашнення овець.
Упродовж останнього десятиріччя під час з’ясування питань доместикації овець стали використовувати більш глибокі біологічні методи, конкретні дані про генетичну мінливість диких баранів і свійських овець.
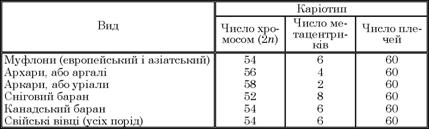
Одним із стійких генетичних показників видових особливостей тварин є хромосомний набір, або каріотип. Нові дані, отримані закордонними і вітчизняними вченими (Г. Манна, М. Тал- кер, 1965; І. Шмітт, Б. Улбріх, 1968; С. Налдер, 1971, 1973; М.В. Орлов, 1975), вказують на відмінності хромосомних наборів диких баранів. Теперішні дикі барани мають 4 форми хромосомних наборів: 52, 54, 56, 58. Нині досліджено хромосомні набори більш як 20 порід свійських овець (Н.С. Бутарін, 1935; І. Маландер, 1959; Р. Борланд, 1964; А. Брюєр, 19б9; Г. Жан- басов, 1975; І.М. Орлов та ін., 1980, К. Шарипов та ін., 1980) і встановлено, що всі вони без винятку мають 54 хромосоми (табл. 12).
Усі дослідники вказують на повну ідентичність європейських і азіатських муфлонів, на збіг одноплечих і двоплечих хромосом і на те, що людина вперше одомашнила тільки муфлонів Малої Азії та Середземномор’я, тобто європейських і азіатських. На думку вчених, усі різновиди диких баранів (аркари, уріали, снігові барани) не є родоначальниками свійських овець. Вівчарство Азії розвивалося на основі використання вже одомашнених овець, завезених із Південно-Західної Азії.
|
Таблиця 1.2. Каріотипи різних видів диких баранів і свійських овець
|
Такої самої думки нині дотримуються і зоологи-систематики (в останньому виданні «Каталогу ссавців СРСР», Ленінград, 1981, вказано, що предками свійських овець були муфлони).
Відтак районами первинної доместикації диких баранів і розвитку культурного вівчарства слід вважати Південну Європу, Передню та Малу Азію. Ця думка підтверджується і тим, що культурні вівці з однорідним руном були створені у давніх цивілізаціях, місця розміщення яких збігаються з ареалом поширення муфлонів.
П.М. Кулешов (1925) зазначав, що більш вірогідно, що довгохвоста вівця була виведена в Сирії, Вірменії і Персії і що всі культурні породи Азії та Європи, в тому числі мериноси цигайські, походять від сирійської вівці.
Відомий дослідник походження тварин С.М. Боголюбський (1959) писав, що створення предків доцигайських овець відбувалося в різних місцях Західної Азії, Кавказу, на південь від Арарату, у верхів’ях Тигру, Євфрату і далі на південний захід у межах Месопотамії та Західного Ірану.
На деяких розкопках матеріальної культури Давнього Вавилону, Урарту є зображення овець із довгою хвилястою вовною.
Н.Б. Янковська (1959) на основі досліджень матеріалів ассирійських колоній III тис. до н.е. відмічає, що серед різних виробів, які поставлялися із різних областей Малої Азії, на другому місці після міді були шерстяні тканини і фарбована вовна.
Все це свідчить, що у давніх овець Малої Азії було однорідне біле руно. Білу благородну вовну мають і нині поширені тут породи овець (балхаська в Туреччині та імеретинська в Грузії, білу- джинська, ідубар в Ірані). Безперечно, що предками всіх цих овець були азіатські муфлони. Білий колір вовни більшості місцевих овець Малої Азії і Південної Європи, очевидно, можна пояснити наявністю у муфлонів білих сідлоподібних по боках на загальному рудо-бурому фоні плям волосяного покриву. Мабуть, ця особливість зумовила відмінності в генетиці забарвлення муфлонів та інших диких баранів. Це добре виявляється при міжвидовій гібридизації свійських овець з білою вовною і диких баранів.
За даними К. Шарипова та ін. (1980), усі нащадки при гібридизації архарів і уріалів з білими матками казахської тонкорунної породи мали коричневе забарвлення вовни. Автори дійшли висновку, що, вірогідно, ген забарвлення диких баранів домінує над білим забарвленням вовни свійських овець.
У дослідах М.Ф. Іванова і Т.П. Белехова (1929) із 13 гібридів, отриманих від спарювання європейського муфлона зі свійською мериносовою вівцею, один був зовсім білий, 9 — білі з невеликими рудими плямами на ногах, холці і шиї і 3 — темно-бурі. До 5-місячного віку руді плями знебарвилися і гібриди стали зовсім білими. Отже, із 13 гібридів було 10 білих і 3 темно-бурих.
При гібридизації азіатських муфлонів з вівцями в типі кори- дель отримали 4 гібриди, із яких при народженні 3 мали зовсім білий окрас, а в одного були світло-коричневі плями на холці, голові та ногах, які потім освітлилися.
Отже, гени пігментації як муфлонів, так і більшості культурних європейських і малоазійських порід овець є рецесивними стосовно білого окрасу, а гени архарів, уріалів, середньоазійських і центральноазійських порід домінантні до білого окрасу.
Одомашнені в Передній і Малій Азії вівці поширилися потім у Середній і Центральній Азії. Не виключено, що на перших етапах одомашнення тварин і переселення людей, коли на гірських пасовищах було багато диких баранів, а свійські ще мало чим відрізнялися від них, дуже часто дикі барани потрапляли у стада свійських овець, відбувалася гібридизація. Тим самим до генофонду свійських овець додавався генофонд архарів або уріалів — так утворювалися нові різновиди тварин. Випадковий характер таких зустрічей і презиготна селекція гамет в напрямі каріотипу свійських овець у більшості поколінь не могли привести до змін у хромосомному наборі тварин, проте це впливало на характер спадковості деяких ознак і властивостей (розмір і окрас волосяного покриву овець).
Потім умови розведення овець і потреби людини поступово примусили вести відбір за м’ясністю, вовновістю, смушковістю, які в результаті роботи упродовж тисячоліть закріпилися у фенотипі різних порід, але не зачепили хромосомний апарат. Отже, в результаті одомашнення овець у Малій і Передній Азії на всьому Азійському континенті було створено численні породи жирнохвостих, курдючних і смушкових порід овець.
Все ж треба зазначити, що поширені в різних регіонах Земної кулі дикі барани дуже мало вивчені біологічною наукою. Всі їх види і раси є безцінним еволюційним даром людства, цілеспрямоване використання якого може багато чого розкрити в домести- каційному і породотворному процесах. Досягнення сучасної біологічної науки дають змогу не тільки збагатити генофонд свійських овець, а й під постійним генетичним та імуногенетичним контролем проводити роботу з «конструювання» нових генотипів, створення свійських порід овець з новим числом хромосом. Ці роботи мають незаперечний не тільки теоретичний, а й практичний інтерес. Велику користь процесові породотворення може дати генофонд муфлонів — у тонкорунному і напівтонкорунному вівчарстві всіх континентів, в романівському вівчарстві, генофонд архарів та уріалів — у м’ясо-сальному і смушковому вівчарстві, генофонд снігових і канадських баранів — у вівчарстві Сибіру, Далекого Сходу та Півночі.