4.1. М’язова тканина
М’язова тканина — це частина м’яса, що має найбільшу поживну цінність. Вона є сукупністю м’язових волокон і сполучнотканинних оболонок, що кількісно переважають.
За морфологічною будовою розрізняють посмуговану мускулатуру, до якої належать скелетні м’язи, і гладку, що входить до складу тканин травного каналу, діафрагми, кровоносних судин, матки та ін.
Мускулатурою змішаного типу є серцевий м’яз.
Найбільший інтерес у технології становить посмугована скелетна мускулатура.
Основним морфологічним і функціональним тканинним елементом посмугованої мускулатури є м’язове волокно.
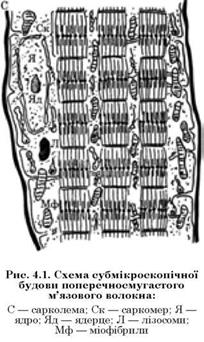 М’язові волокна — це своєрідна гігантська багатоядерна клітина завтовшки від 10 до 100 мкм і завдовжки 12 см і більше. Структура м’язового волокна дуже складна. Поверхня його вкрита еластиновою оболонкою — сарколемою (рис. 4.1).
М’язові волокна — це своєрідна гігантська багатоядерна клітина завтовшки від 10 до 100 мкм і завдовжки 12 см і більше. Структура м’язового волокна дуже складна. Поверхня його вкрита еластиновою оболонкою — сарколемою (рис. 4.1).
Крім сарколеми, в структурі м’язового волокна можна розрізнити тонкі поздовжні структури — міофібрили, а також ядра і кілька органел: мітохондрії, рибосоми, лізосоми та ін. Меншу частину клітин (35 — 40 %) становить саркоплазма, що повністю оточує всі інші утворення.
М’язові волокна складаються в первинні м’язові пучки. У пучках волокна з’єднані найтонши- ми прошарками сполучної тканини, зв’язаними з волокнами ендомізієм. Ендомізій утворюється тонкими і ніжними колагеновими й еластиновими волокнами, зібраними в пучки, вільний простір між ними заповнений проміжною речовиною.
Первинні м’язові пучки об’єднуються в пучки вторинні і т.д. Пучки вищого порядку вкриті міцнішою сполучнотканинною оболонкою — перемізієм і в сукупності складають м’яз. Ендомізій і перемізій утворюють своєрідний каркас або строму м’язів. їх міцність впливає на жорсткість м’язової тканини. Тому на практиці їх виділяють в окрему категорію внутрішньом’язової сполучної тканини.
М’яз також вкритий оболонкою — епімізієм. Перемізій і епімі- зій побудовані з колагенових волокон різної структури і міцності, що утворюють більш-менш складні сплетіння і вміщують різну кількість еластинових волокон. У перемізії й епімізії м’язів деяких видів відгодованих тварин є жирові клітини, що утворюють так звану «мармуровість м’яса» на його поперечному розрізі.
Сарколема побудована з двох шарів з ліпідним прошарком. Внутрішній (плазматичний) шар має вигляд мембрани завтовшки 0,1 мкм; зовнішній — базальний шар, складається з густої сітки волокон двох видів: переважно колагенових (діаметром близько 0,2 нм) і безструктурних еластинових.
Сарколема має вибіркову проникність. За життя тварин через неї проходять складові молекул вуглеводів, жирних кислот, амінокислот, білків. їх переміщення залежить від осмотичного тиску і активної регуляції сарколеми. її проникність регулюється нервовою системою і змінюється під час роботи м’язів, пов’язаної з під- кисненням вмісту волокна.
Міофібрили є активними скорочувальними елементами м’язового волокна, що займають більшу частину клітини (близько 60 — 65 %). Це волокнисті посмуговані структури (завтовшки 1 — 2 нм), які розміщені в протоплазмі м’язового волокна поздовжньо направленими паралельними пучками.
Під мікроскопом за великого збільшення можна побачити ділянки, що рівномірно чергуються, одні з яких у прохідному світлі здаються темними, інші — світлішими.
При дослідженні в поляризаційному мікроскопі темні ділянки міофібрил виявляють подвійне променезаломлення (анізотропні диски, або диски А), а світлі не мають таких властивостей (ізотропні диски, або диски Л). Така оптична неоднорідність цих ділянок зумовлена неоднаковою їх будовою та білковим складом (рис. 4.2).
|
Рис. 4.2. Схема будови саркомера у розслабленому стані: БМС — базальна мембрана сарколеми; ПМС — плазматична мембрана сарколеми; А — анізотропний диск; J — ізотропний диск; Н — світлий диск; М — смужка; Z — пластина; а — активні протофібрили; м — міозинові протофібрили; ТС — канали Т-системи; БТ — бокові тяжі; Тр — тріади саркоплазматичного ретикулу- ма; Пср — поздовжні канали саркоплазматичного ретикулума |
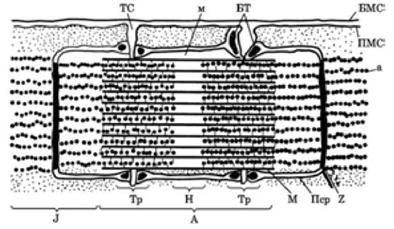
Основні ділянки міофібрил, обмежені мембранами Z, називають саркомером. Довжина саркомера 2 — 3 нм. Кожна міофібрила складається з кількох саркомерів.
Кожну міофібрилу можна розщепити (наприклад, за допомогою ультразвуку) на численні найтонші нитки, які видно тільки під електронним мікроскопом. Ці елементарні волоконця, що складають міофібрили, називають протофібрилами. Розрізняють товсті й тонкі протофібрили (нитки). Товсті нитки (діаметром близько 100 нм і завдовжки 450 нм) побудовані з білка міозину, розташовані паралельними рядами в диску А. Посередині диска А в зоні Н товсті нитки збільшуються в діаметрі, утворюючи зону м (див. рис. 4.2).
Тонкі нитки (діаметром близько 58 нм), що складаються з білка актину, ідуть від лінії Z через диск ^ їхні кінці знаходяться між товстими нитками в зоні А. У зоні Н тонких ниток немає. Товсті й тонкі нитки з’єднані перемичками (містками) завтовшки приблизно 30 нм.
Будова тонких і товстих ниток доволі складна. Товсті складаються з паралельно розташованих, стиснених молекул міозину (кожна нитка, що входить у диск А, вміщує від 200 до 400 молекул білка). Через певні проміжки (429 і 143 нм) уздовж нитки розміщені містки, що стикаються з містками на тонких нитках. Вони ніби закручені навколо спіралі і утворюють піки, подібні до зубців (ніжок). Тонкі нитки складаються з двох спірально переплетених ланцюгів молекулярного актину (побудованих з мономерного актину). Один ланцюг закручений навколо іншого, осьового.
На поперечному розрізі міофібрил товсті й тонкі нитки орієнтовані так, що утворюють гексагональну структуру. Фібрили оточені й тісно пов’язані з особливою структурою, що складається з трубочок та бульбашок і називається саркоплазматичним ретикулумом.
Саркоплазма. У саркоплазмі потрібно розрізняти шар зернистої цитоплазми, що прилягає до ядер, і міжфібрилярну плазму, що заповнює проміжки між фібрилами.
Зерниста цитоплазма вміщує різні органоїди (організовані внутрішньоклітинні утворення, які беруть участь в основних життєвих функціях).
Ядра розташовані по периферії клітини під оболонкою. Вони мають овальну, плоску форму. Структура ядер неоднорідна. В них вміщуються переважно дезоксирибонуклеопротеїни. Ядра вкриті надтонкою оболонкою, що складається з двох шарів білкових молекул, між якими лежить ліпідний прошарок.
Мітохондрії — це утворення кулястої або видовженої форми діаметром 0,7 — 1,0 мкм (2 — 7 мкм). Вони обмежені двошаровою мембраною завтовшки до 180 нм, що складається з глобулярних білкових молекул та шару молекул ліпідів. У середині мітохондрії розташовані також двошарові мембрани, що складаються з білко- во-фосфоліпідних комплексів у вигляді перегородок, гребенів.
Рибосоми (саркосоми) — це утворення круглої або овальної форми діаметром від 200 до 300 нм. Вони складаються переважно
з білкової оболонки і рибонуклеїнових кислот (РНК).
Лізосоми — нестійкі утворення сферичної форми (середній діаметр близько 0,4 мкм), оточені ліпопротеїдною мембраною. У структурі локалізовані різні гідролітичні ферменти. Фізіологічною функцією лізосомів є внутрішньоклітинне перетравлення.
М’язова тканина характеризується складним хімічним складом. До неї входить значна кількість побічних речовин, вміст і властивості яких можуть змінюватися залежно від багатьох факторів як за життя тварин (передзабійне утримання), так і відразу після забою.
Масова частка основних хімічних речовин у м’язовій тканині (препарованій) ссавців становить, %:
|
|

Вміст води у м’язах коливається залежно від віку тварин: чим вони молодші, тим більше вологи в м’ясі. Вміст води в різних групах м’язів неоднаковий і зменшується у міру збільшення вмісту жиру.
Після висушування м’язової тканини сухий залишок становить близько 30 %, у тому числі органічних речовин 20 — 28 %; неорганічних солей — 1,0 — 1,5 %.
Основним компонентом органічних речовин тканини є білки. На їхню частку припадає близько 80 % сухого залишку або 16,5 — 20,9 % маси тканини. Чільне місце належить численним екстрактивним речовинам (азотистим і безазотистим), що виконують важливу роль у біохімічних перетвореннях м’язів. Деякі з них є проміжними або кінцевими продуктами обміну. До складу сухого залишку входять також жири й інші ліпіди. У м’язовій тканині містяться різні вітаміни.