4. Окислювально-відновний потенціал
Протягом десятиліть було відомо, що мікроорганізми проявляють мінливу чутливість до окислювально-відновного (ОВ) потенціалу (O/R, Eh) середовища перебування. Як правило, ОВ потенціал субстрату може бути визначений як легкість, з якої субстрат втрачає або здобуває електрони. Коли елемент або речовина втрачає електрони, субстрат окисляється, тоді як субстрат, що отримує електрони, стає відновленим: ![]() .
.
Окислювання може також бути досягнуто додаванням кисню, як показано на наступній реакції: ![]() .
.
Тому речовина, що легко віддає електрони, - чудовий відновлювач, а та, що легко приймає електрони, - гарний окислювач. Коли електрони переносяться від однієї речовини до іншої, між цими речовинами створюється різниця потенціалів. Ця різниця може бути виміряна при використанні підходящого устаткування й виражена в мілівольтах (мВ). Чим більше окислена речовина, тим більше позитивним буде його електричний потенціал. Коли концентрації окислювачів і відновлювачів рівні, електричний потенціал дорівнює нулю. ОВ потенціал системи позначається символом Eh. Для росту аеробних мікроорганізмів необхідні позитивні значення Eh (окислені), тоді як для анаеробних – негативні значення Eh (відновлені) (рис. 15). Серед речовин у харчових продуктах, які допомагають підтримувати відновлені умови, можна виділити SH-групи в м'ясі й аскорбіновій кислоті й цукри, що редукують, у фруктах й овочах. Що стосується максимальних позитивного й негативного значень мВ на рис. 15, вони не тільки необхідні для росту аеробів й анаеробів, але ці граничні рівні можуть також бути летальними для відповідних груп.
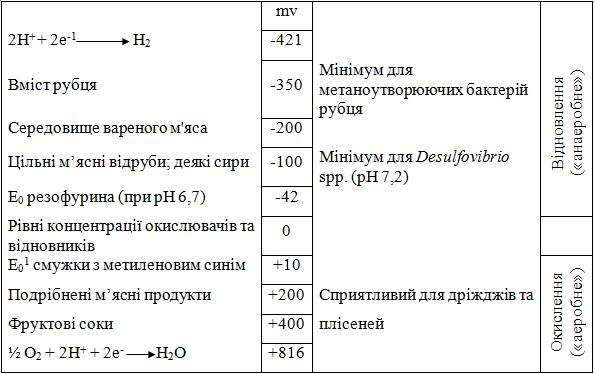
Рис. 15. Схематичне зображення окислювально-відновних потенціалів щодо росту певних мікроорганізмів.
ОВ потенціал харчових продуктів визначається в такий спосіб:
1. Специфічний ОВ потенціал сировини.
2. Буферна ємність – це опірність зміні потенціалу харчових продуктів.
3. Тиск кисню атмосфери навколо харчового продукту.
4. Доступність, що атмосфера має до продукту.
Що стосується потреби мікроорганізмів у певному ОВ потенціалі, деяким бактеріям для початку росту необхідні відновлені умови (Eh приблизно − 200 мВ), тоді як іншим бактеріям потрібний для росту позитивний Eh. До першої категорії належать анаеробні бактерії, такі як рід Clostridium; до другого належать аеробні бактерії, такі як деякі члени роду Bacillus. Деякі аеробні бактерії в дійсності ростуть краще при злегка відновлених умовах, і ці організми відносяться до мікроаерофілів. Приклади мікроаерофільних бактерій: лактобацили й кампілобактери. Деякі бактерії здатні рости як при аеробних, так і при анаеробних умовах. Такий тип бактерій називається факультативні анаероби. Більшість плісеней і дріжджів, що виділяють з поверхні продуктів і зсередини, - аероби, хоча невелика кількість має риси факультативних анаеробів.
Що стосується ОВ потенціалу рослинних харчових продуктів, особливо рослинного соку, то вони мають значення Eh від +300 до +400 мВ. Не дивно, що аеробні бактерії й цвілі найчастіше викликають псування продуктів цього типу. Цільні м'ясні продукти мають значення Eh біля –200 мВ; у фарші Eh звичайно дорівнює +200 мВ. Сири різних типів мають ОВ потенціал у негативній області: від -20 до -200 мВ.
Що стосується ОВ потенціалу м'язів до настання посмертного задубіння на противагу м'язам після посмертного задубіння досліджували роботу з виміру Eh м'язів протягом 30 годин після забою та встановленню його впливу на ріст анаеробних бактерій. Ці автори виявили, що ОВ потенціал м'яза stemocephalicus коня відразу після смерті дорівнював +250 мВ, у цей час клостридії втратили здатність розмножуватися. Через 30 год після смерті ОВ потенціал упав приблизно до 30 мВ у відсутності бактеріального росту. Коли з'явилася можливість для бактеріального росту, Eh упав до -250 мВ. Ріст клостридій спостерігався при значенні ОВ потенціалу 36 мВ і нижче. Ці автори підтвердили, що дані для м'яса коней характерні й для м'яса китів: анаеробні бактерії не розмножуються до настання посмертного задубіння, тому що в м'яса до цього моменту високий ОВ потенціал. Подібне безперечно вірно й для яловичини, свинини й інших видів м'яса.
Мікроорганізми впливають на ОВ потенціал свого навколишнього середовища під час росту, змінюючи значення рН. Це вірно, особливо для аеробів, які можуть знижувати ОВ потенціал свого середовища, тоді як анаероби не можуть. При рості аеробів О2 у середовищі знижується, приводячи до зниження ОВ потенціалу. Ріст не сповільнюється доти, поки клітини здатні використати О2-донуючі або водень-акцептуючі речовини із середовища. У результаті середовище стає бідніше окисленими й багатше відновленими субстратами. ОВ потенціал середовища може бути знижений мікроорганізмами за допомогою синтезу ними певних побічних продуктів метаболізму таких, як H2S, що здатний знижувати ОВ потенціал до −300 мВ. Оскільки H2S швидко вступає в реакцію із О2, він буде накопичуватися тільки в анаеробних умовах.
ОВ потенціал залежить від показника рН субстрату, і прямий взаємозв'язок між двома факторами – це величина r, обумовлена в такий спосіб:
Eh=2,303(R*T/F) * (rH-2pН) (4)
де R=8,315 Дж, F=96500 Кл, а T – абсолютна температура.
Отже, значення рН субстрату повинне встановлюватися, коли заданий ОВ потенціал. Звичайно Eh установлюється при рН 7,0 (позначається Eh'). У випадку, якщо ОВ потенціал установлюється при рН 7,0, 25 ºС і з сумарною концентрацією всіх речовин 1,0 M, то Eh=Eh'0 (спрощене рівняння Нернста). У природі ОВ потенціал прагне бути більш негативним при прогресивно лужних умовах.
Серед поживних речовин, що зустрічаються в природі, аскорбінова кислота й цукри, що редукують, у рослинах і фруктах й -SH групи в м'ясних продуктах мають першорядне значення. Присутність або відсутність підходящої кількості окислювально-відновних агентів у середовищі має очевидну важливість для росту й активності мікроорганізмів.
У той час як ріст анаеробів, як звичайно вважають, має місце при знижених значеннях ОВ потенціалу, виключення кисню може бути необхідним для деяких анаеробів. Коли Clostridium perfringens, Bacteroides fragilis й Peptococcus magnus культивувалися в присутності О2, гальмування росту спостерігалося навіть у тому випадку, коли середовище мало негативний Eh −50 мВ. Ці дослідники виявили, що ріст відбувався в середовищі з високим ОВ потенціалом 325 мВ у відсутності кисню.
Що стосується впливу ОВ потенціалу на синтез ліпідів Saccharomyces cerevisiae, було показано, що зростаючі в анаеробних умовах клітини виробляють більш низьку сумарну кількість ліпідів, сильно варійовану гліцеридну фракцію й знижені кількості фосфоліпідних і стеролових компонентів у порівнянні з аеробно зростаючими клітинами. Ліпіди, що продукуються анаеробно зростаючими клітинами, характеризуються високим вмістом (аж до 50% сумарних кислот) 8:0 й 14:0 кислот і низьким рівнем ненасичених жирних кислот у фосфоліпідній фракції. В аеробно зростаючих клітинах 80…90% жирнокислотних компонентів було зв'язано в гліцериди, а фосфоліпіди, як було виявлено, були представлені 16:1 й 18:1 кислотами. На відміну від аеробно зростаючих клітин, анаеробно зростаючі клітини S. cerevisiae мали потреби в ліпідах і стеролі.