1.2.2.2. БІОЛОГІЧНІ ОСОБЛИВОСТІ ПОЛЬОВИХ КУЛЬТУР
До біологічних особливостей відносять способи розмноження, морфологічну будову та структуру, реакцію на середовище і добрива, температуру проростання насіння, здатність до відростання (отавність), ріст і розвиток рослин, особливості життєвого циклу, тривалість періоду вегетації, будову, потужність та вбирну здатність кореневої системи, особливості наливу і достигання зерна, врожайність, її структуру та ін.
Способи розмноження. Розрізняють два способи розмноження рослин — генеративний (насінням) і вегетативний (бульбами, частинами кореневищ). У рослинництві застосовують переважно генеративний спосіб. Вегетативним способом розмножують і вирощують картоплю і топінамбур. Застосовують його і в травосіянні, зокрема при вирощуванні кореневищних злаків — стоколосу безостого, пирію повзучого, конюшини білої (повзучої). Після поверхневого обробітку ці злакові трави розмножують відростанням з частинок кореневищ. Повзучі пагони конюшини білої вкорінюються і утворюють нові рослини.
Останнім часом широко застосовується новий метод вегетативного розмноження — мікроклонування — вегетативне відновлення рослин з меристемних клітин. Рослини, одержані таким чином, ділять на відрізки і багаторазово вегетативно розмножують на спеціальних поживних розчинах до висаджування в ґрунт. Цей метод широко застосовують у насінництві картоплі.
Невеличкі рослинки — зародки цукрових буряків, одержані з клітин, можна використовувати як вегетативний насіннєвий матеріал замість звичайного насіння, правда, переважно у селекційній роботі, оскільки технологія одержання такого насіннєвого матеріалу поки що дуже дорога.
Ріст рослин і його регулювання. У загальних рисах ріст рослин — це переважно кількісні зміни в рослині, спрямовані здебільшого на збільшення її маси, на відміну від розвитку, пов'язаного з якісними змінами в рослині у процесі її онтогенезу (повного циклу розвитку, починаючи із запліднення — зиготи). Є різні тлумачення поняття росту. На думку Д. А. Сабініна (1940, 1963), наприклад, ріст — це процес новоутворення елементів структури організму (органів, клітин, їх частин і навіть субмікроскопічних компонентів протоплазматичних структур до макромолекул включно). Це визначення, звичайно, не завжди відповідає поняттю росту в практиці агрономії і науковій роботі — тут це поняття трактується ширше за уявлення про нього як лише про збільшення маси та об'єму рослин і їх органів з урахуванням елементів росту, коли не спостерігається збільшення маси рослин, а відбувається утворення (диференціювання і нагромадження) її органів («прихований ріст»). Може спостерігатись навіть зменшення маси рослин (рослина «худне»). Це зумовлюється несприятливим періодом вегетації, створенням генеративних органів та ін. Так, у кукурудзи в період викидання волотей маса рослин не збільшується, а навіть зменшується. Такі зміни динаміки нагромадження маси свідчать не про відсутність росту, а лише про його характер (Д. А. Сабінін).
Відомий учений-біолог В. С. Шевелуха (1992) у своїй праці «Ріст рослин і його регуляція в онтогенезі» визначає ріст як процес диференціювання структури організму за рахунок утворення нових і збільшення старих його елементів (молекул, органел, клітин, тканин і органів).
Відростання (кущення) злакових і бобових багаторічних трав також є різновидом вегетативного розмноження. У високоотавних однорічних трав із сплячих бруньок на кореневій шийці, вузлі кущення, у пазухах листків нижніх частин стебел з'являються пагони «сисунці». Залежно від умов вегетації (переважно умов світлового режиму) вони можуть бути генеративними (плодоносними) або вегетативними.
При поліпшенні агрофізичних властивостей ґрунту та поєднанні вегетативного розмноження багаторічних трав з періодичним їх обсіменінням травостої можна експлуатувати десятки років.
Отже, вегетативне розмноження в рослинництві та його поєднання з генеративним розмноженням можна застосовувати для вирощування багатьох сільськогосподарських культур. Однак основним у розмноженні абсолютної більшості польових культур є генеративний спосіб.
Насіннєва продуктивність різних польових культур неоднакова. Тобто культури мають різні коефіцієнти розмноження. Високі вони у пшениці, кукурудзи, сорго, соняшнику, ріпаку, суданської трави, могару, гарбузів, кавунів, коренеплодів. Водночас зернобобові (горох, боби, соя, люпин білий і жовтий) і багаторічні трави (люцерна, конюшина, еспарцет) мають незначні коефіцієнти розмноження (10 — 15). Невисокий коефіцієнт вегетативного розмноження і в картоплі (8 - 12).
Для початку вегетації або її відновлення зерно, бульби, вузли кущення, кореневі шийки, нижні частини стебел, підземні стебла повинні мати певний запас поживних речовин. Щоб ці речовини раціонально використовувалися в період проростання насіння, треба забезпечити необхідні умови: оптимальні строки сівби культури, відповідні глибину загортання насіння, вологість і температуру ґрунту та ін. При знижених температурах або недостатній вологості ґрунту розтягуються строки появи сходів, відростання бруньок запасу, витрачається надмірна кількість поживних речовин і, як результат, зріджуються сходи або травостій, погіршується ріст рослин.
Відростання (отавність) польових рослин. Деякі культури можуть відростати після скошування. Певною мірою цю властивість мають озимі і ярі хліба (коли вони використовуються на корм), кукурудза, сорго зернове. Найбільша отавність у суданської трави, пажитниці багатоквіткової, пайзи, конюшини олександрійської, персидської, підземної, лучної (червоної), рожевої і білої, люцерни посівної і жовтої, еспарцету піщаного і закавказького, лядвенцю рогатого, грястиці збірної; добра — у вівсяниці лучної і очеретяної, тимофіївки лучної, райграсу високого, стоколосу безостого. Задовільна отавність у сорго, ріпаку озимого, еспарцету посівного (виколистого), пирію без-кореневищного, буркуну дворічного, серадели; досить низька — у вівса, вики ярої, часто озимої і паннонської, жита кормового, могару, буркуну однорічного.
Злакові культури відростають із сплячих бруньок вузла кущення, бобові, хрестоцвіті та інші стрижнекореневі — з кореневої шийки і пазушних бруньок нижньої частини стебла (на висоті до 8 — 10 см, іноді вище).
Отавність значною мірою залежить від умов зволоження, фази збирання, висоти зрізу при скошуванні. Певне значення може мати і спосіб збирання. Так, застосування роторних косарок при збиранні люцерни, конюшини і еспарцету погіршує їх відростання, оскільки стерня розщеплюється до кореневої шийки рослин, залишки стебел з пазушними бруньками підсихають. Позитивно впливають на відростання внесення азотних і калійних добрив, аерація ґрунту (коли 75 — 80 % ґрунтових пор займає вода, а решту — повітря), що справляє вирішальний вплив на розподіл, перерозподіл і використання створених у процесі фотосинтезу та метаболізму органічних речовин, а також поглинутих мінеральних солей і води, які йдуть на утворення нових органів і тканин, їх регенерацію та на запасні відкладення. У визначенні В. С. Шевелухою суті росту організмів практично збережена головна ідея Д. А. Сабініна про ріст як процес самоутворення елементів структури організму. Проте в його формулюванні підкреслюється потреба в більш активному вивченні росту як своєрідного біологічного рушія, який мобілізує сили рослинного організму, і як атрагуючого (притягального) регулятора, котрий посилює або послаблює процеси утворення, обміну й руху речовин в організмі.
Особливості росту польових рослин. Виділяють такі особливості росту рослин: інтегральність, що характеризується і описується S-подібною кривою; параболічний, який стосується об'єктів і систем і визначає основну їх якість і особливість. Розрізняють також генетичну, гормональну й екологічну зумовленість росту, його інтенсивність, масштабність, просторову локалізацію. Всі ці характеристики залежать від внутрішніх факторів розвитку та умов вегетації рослин.
Ріст рослин характеризується високою чутливістю ростових процесів до зміни погоди, зокрема фізіолого-біохімічних і біоелектричних потенціалів у рослині, тобто адаптивними її можливостями, які необхідно враховувати при оптимізації процесу вирощування.
Ріст рослин характеризується також лабільністю (рухомістю) основних показників, яка зумовлена високою чутливістю до зміни зовнішніх і внутрішніх факторів вегетації.
У процесі росту рослини пристосовуються до зміни умов вегетації. Цьому сприяють спадкові біофізичні та біохімічні особливості клітин, які забезпечують життєдіяльність організму, в тому числі ріст у широких для кожного виду рослин межах температурних, світлових та інших умов.
Певна локалізація і послідовність ростових процесів у часі й просторі визначаються так званим каскадним характером генної регуляції і спостерігаються в рослинах, рослинних угрупованнях — ценозах усіх кліматичних зон, зумовлені спадковою програмою організму, поєднанням зовнішніх і внутрішніх факторів вегетації рослин, кліматичними особливостями зон та географічних поясів.
У процесі росту рослин відбувається його саморегуляція. За даними В. С. Шевелухи, вона чітко виявляється у віковій мінливості та її масштабності, інтенсивності й локалізації, в законі великого періоду росту органів, в ендогенній добовій і пульсуючій періодичності та ритмічності, здатності рослин швидко відновлювати рівень ростових процесів, порушений коливанням різних факторів вегетації. Така саморегуляція є найважливішою особливістю всіх біологічних об'єктів.
Характерними для росту є нерівномірний хід у часі, періодичність, ритмічність. Це виявляється у закономірних або випадкових змінах показників інтенсивності росту в часі. Вони генетично зумовлені в процесі еволюції і пов'язані із сезонними, добовими, погодинними та мікрочасовими періодами вегетації.
Екзогенні коливання росту регулюються зовнішніми факторами середовища, ендогенні — «біологічним годинником», інтенсивністю нуклеїнового і білкового синтезу, темпами утворення, нагромадження і активності ферментних та ізоферментних систем, фітогор-монами й іншими продуктами метаболізму рослин (В. С. Шевелуха, 1992).
Регуляторна функція росту в онто- й морфогенезі рослин виявляється у його впливі на швидкість і напрям метаболічних процесів синтезу, розпаду, руху і нагромадження органічних сполук та інших речовин, у їх розподілі та реутилізації під впливом атрагуючої дії ростучих органів рослин —могутніх центрів поглинання речовин.
Для росту рослин, як і інших біологічних об'єктів, властивий високий ступінь кореляції (взаємозв'язку) його показників з параметрами продукційного процесу і ходу формування врожаю. Ця особливість дає змогу використовувати її як тест під час агрономічного контролю і програмування врожаю.
Ці особливості ростових процесів, які по суті вперше сформулював В. С. Шевелуха, є найбільш загальними для рослин і виявляються на всіх рівнях організації біологічних систем — молекулярному, клітинному, організмовому, популяційно-видовому та біосфер-ному. Отже, вони є загальними, оскільки відображують загальні закономірності росту біологічних об'єктів, тому безпосередньо стосуються особливостей вегетації польових культур.
Добова періодичність росту польових культур. Ріст у його практичному значенні визначають за його лінійними показниками у процесі вегетації. При спостереженні за ростом бульбоплодів беруть проби по фазах або по декадах. Результати таких спостережень дають загальне уявлення про розміри рослин, коренів, бульб, динаміку формування врожаю. При цьому можливі різні залежності між лінійним ростом культури і врожаєм при неоднаковій щільності травостою. Приріст рослин можна визначити за рівняннями регресії. Наприклад, залежність між приростом трави грястиці збірної і відносною швидкістю лінійного росту її листків та пагонів у відсотках до попередньої швидкості можна виразити такими рівняннями регресії (за В. С. Шевелухою):
Y1 = -131,1 + 2,57x —при щільності травостою до 1800 пагонів на 1 м2; Y2 = -464,2 + 6,36x — при щільності до 3000 пагонів на 1 м2; Y3 = -535,5 + 6,78x — при щільності до 3500 пагонів на 1 м2,
де 1і_з — приріст врожаю за одиницю часу, % до початкового; х —
відносна швидкість лінійного росту листя і пагонів, % до початкової.
Між показниками росту бадилля картоплі в період від сходів до середини цвітіння і сухою масою бадилля існує корелятивний зв'язок (r= 0,88 ±0,15 і r = 0,98±0,08), а кореляція між цими показниками і бульбами є значно нижчою: r = 0,69 ± 0,32 і r = 0,53 ± 0,37.
Виявлено також високу залежність росту коренеплодів уночі від швидкості їх росту в денні часи (r= 0,640-0,775). Отже, ріст рослин може характеризуватись математичними показниками залежності між умовами вегетації, ростом вегетативних органів, коренів, бульб, формуванням зерна та ін.
Для загальної характеристики формування врожаю рослин важливим є врахування добової, або циркадної, періодичності росту польових культур. Циркадний ріст— це ритм або циклічні коливання інтенсивності біологічних процесів, в тому числі лінійних показників, приблизно за добу (20 - 28 год). Вони мають велике значення для пізнання загальних закономірностей росту рослин. Ці дані отримують за допомогою спеціальних приладів — ауксанометрів, які фіксують наявність незначних приростів листків, стебел, бульб. В. С. Шевелуха виділяє 10 типів циркадного росту рослин.
1. Синусоїдальний (наближено синусоїдальний) тип росту рослин можна зобразити у вигляді синусоїди або наближеної до неї за формою кривої з фазами максимуму й мінімуму в ранні ранкові години доби. Фаза мінімального росту рослин збігається з мінімальними температурами і виявляється чітко в часі — рано вранці (4 — 6 год), максимум — залежно від чутливості рослин до вологи, температури, світла, тобто залежно від екологічних особливостей рослин може припадати на 10 - 12, 14 - 16, 17 - 19 год, а весь період має циркад-ну тривалість.
Цей тип ростових процесів спостерігається у злакових зернових і трав, а в таких культур, як льон, коноплі, кормові боби, амплітуда коливань менша, ніж у злаків. Близький до синусоїдального добовий ріст бобових трав, люцерни, еспарцету, лядвенцю рогатого.
Кутовий тип росту рослин має вигляд добової кривої під різ
ним (гострим або тупим) кутом з однією фазою максимуму й міні
муму. Як і за першого типу росту, фаза мінімуму припадає на ран
ній ранок, а швидкість денного росту залежить від позитивних по
казників основних екологічних факторів. Оптимум їх припадає на
17 — 20 год, тобто під вечір. Цей тип росту спостерігається у люпину
жовтого. Амплітуда коливання росту цього типу в 1,5 раза нижча
порівняно з першим і становить 2 — 3 мм/год.
2. Параболічний тип росту спостерігається у коренеплідних рос
лин, зокрема у брукви. Найкраще вони ростуть у вечірні години,
тобто коли настає прохолода (20 — 21 год). Вдень у сонячну погоду
ріст коренеплодів уповільнюється через зниження відносної волого
сті повітря. Амплітуда коливань росту більша, ніж за кутового типу
росту, але менша, ніж при синусоїдальному, — 4 — 5 мм/год.
3. Імпульсний тип характеризується стрибкоподібним ростом,
причому вранці ріст уповільнюється, а максимум припадає на нічні
години — з 20 — 21 до 7 — 8 год ранку. Цей тип росту характерний
для листків цукрових і кормових буряків.
4. Імпульсний тип росту з позитивними і негативними показни
ками спостерігається в коренеплодів цукрових і кормових буряків,
брукви, бульб картоплі. Він може бути на рівні проростків і на висо
кому рівні. Негативний вплив сонячного світла, певне зневоднення
тканин рослин вдень негативно позначаються на добовому прирості
коренеплодів і бульб картоплі, а тому вони вдень практично не рос
туть.
5. Імпульсно-релаксаційний тип росту з позитивними і негатив
ними показниками характерний для моркви. Імпульсне посилення
ростових процесів спостерігається о 16—17 год. Швидкість росту до
сягає максимуму о 20 — 21 год, з поступовим зниженням до мінімуму
о 7 - 8 год.
6. Імпульсно-релаксаційний тип без негативних показників рос
ту в денний час подібний до попереднього типу росту, але з менш
вираженим пригніченням ростових процесів вдень. За цим типом
відбувається вегетація коренеплодів на другий рік вирощування
цукрових і кормових буряків, моркви. Ростові процеси досить актив
ні — 0,4—1,1 мм/год. Найкраще процеси росту відбуваються при
пониженому освітленні і підвищеній відносній вологості повітря.
Оскільки під час цвітіння рослини потребують підвищеної температури, в нічні години спостерігається релаксація ростових процесів, в ранішні — різке сповільнення росту. Амплітуди коливань приросту досить значні — 0,3 — 0,9 мм/год. За таким типом відбувається добовий ріст конюшини.
8. Релаксаційний тип характеризується рівномірним прискорен
ням росту вдень, увечері і вночі, до 7 — 8 год ранку. Мінімальні його
значення спостерігаються близько 10 год. За таким типом відбува
ється ріст листків моркви.
9. Двохвильовий тип росту спостерігається у стебел і листків кар
топлі, в конюшини лучної. Протягом доби швидкість росту двічі до
сягає максимальних і мінімальних значень. Денний мінімум швид
кості росту спостерігається о 12—15 год, нічний — о 1 — 2 год, мак
симум — о19-21і4-6 год.
10. Відносно рівномірний вирівняний хід росту рослин та їх ор
ганів спостерігається при понижених або різких градієнтах основ
них факторів середовища у злакових, льону, конопель, бобових,
картоплі і коренеплодів. Умови вегетації характеризуються низькою
температурою, яка різко не змінюється. Якщо такі умови тривають
1-2 доби, то криві росту ще нагадують попередні типи, наприклад,
синусоїдний або кутовий. При тривалому впливі низьких добових
градієнтів факторів середовища ритмічності майже не спостеріга
ється. Має місце рівномірний, більш-менш рівний добовий хід
швидкості росту рослин.
Регулювання ростових процесів рослин. Взаємозалежність росту і фізіологічних процесів у рослині (дихання, фотосинтез) не завжди чітка. Кореляція між ростом і диханням більшою мірою спостерігається у злакових і найменшою — у коренеплідних і бульбоплідних рослин. За більш тривалий час (у середньому за кілька діб) прирости мають більшу кореляцію з показниками інтенсивності основних фізіологічних (енергетичних) процесів у рослинах. Важливу роль при цьому відіграє здатність рослин адаптуватися до умов вегетації. Вона значною мірою залежить від спадкових — біологічних і екологічних особливостей рослин — холодостійкості, реакції на відносну вологість повітря, стресові умови (температурні максимуму, висока сонячна інсоляція та ін.). У зв'язку з цим виникає потреба в регулюванні росту і вегетації рослин в цілому проведенням відповідних агротехнічних заходів та застосуванням регуляторів росту (природних і синтетичних), які взаємодіють з фітогормонами. Особливо цінними є так звані антистресові засоби, зокрема картолін (оксикар-бін). Нині є багато груп регуляторів росту й розвитку рослин (гібереліни, ауксини, абсцизова кислота, цитокініни, фузикокцини, етилен та їх синтетичні аналоги), котрі впливають на ріст, морфогене-тичні особливості та інші показники, сприяють підвищенню врожайності. Так, цитокінін позитивно впливає на ростові процеси, зокрема на синтез РНК. Кремнійорганічні сполуки (мівал і крезацин) діють на рослину як біостимулятори, поліпшують обмін речовин, підвищують стійкість (адаптацію) рослин до несприятливих умов, тобто мають антистресову дію, підвищують врожайність. Особливо позитивно впливає на ріст, адаптацію рослин до несприятливих умов, їх урожайність крезацин-тріс (2-оксиметил), амоній-крезоксиацетат або триетаноламінова сіль крезоксиоцтової кислоти. Він нетоксичний, не має мутагенної, канцерогенної, тератогенної і кумулятивної дії.
Проте більшість регуляторів росту рослин (ретардантів) можуть мати певний негативний екологічний вплив. Тому слід постійно контролювати використання цих препаратів. А для регулювання росту, підвищення продуктивності культур і якості продукції підбирати кращі сорти і гібриди, вдосконалювати обробіток ґрунту, догляд за посівами.
Як показали дослідження, проведені в Інституті зернового господарства УААН (А. Г. Мусатов, 1997), для досягнення високого рівня технології слід враховувати гідрометеорологічні умови, їх вплив на вегетацію (ріст) рослин не в цілому, а по фазах вегетації. Те саме стосується умов живлення рослин. Наприклад, для ярих зернових велике значення має оптимізація умов росту рослин у фази кущення, колосіння (викидання волоті), наливу зерна.
Розвиток рослин. Розрізняють стадії і фази процесу вегетації польових культур. Крім того, в житті рослин виділяють періоди та етапи органогенезу, які припадають на певні фази утворення і розвитку органів рослин.
Стадії розвитку відбуваються в проростаючому насінні й точках росту бруньок рослин у період вегетації до настання кущення (пагоноутворення). Вони по суті є внутрішніми біохімічними й фізіологічними змінами рослини, які відбуваються під впливом температури та світла.
Протікання стадії яровизації, світлової та деяких інших фаз є якісним переходом від вегетативного циклу, який супроводжується лише нагромадженням маси рослин, до генеративного. Ознакою того, що в рослини відбулися ці стадії, є перехід її від фази кущення (пагоноутворення) до фази виходу в трубку у злакових і гілкування у бобових, хрестоцвітих та інших стрижнекореневих рослин. Це означає, що в рослині (або стеблах куща) сформуються суцвіття і настане генеративний період.
Фази вегетації. У злакових, бобових і хрестоцвітих та інших культур розрізняють такі фази вегетації: проростання, кущення у злакових; пагоноутворення у бобових, хрестоцвітих та інших стрижнекореневих; вихід у трубку у злакових; гілкування у бобових та ін.; колосіння (або викидання волоті) у злакових; бутонізація, цвітіння, плодоношення в інших культур.
Міжфазний період триває 7-12 днів і залежить переважно від температури. Чим вища температура у фазі, тим коротший міжфазний період.
Початком фази вважається настання її у 10 — 15 % рослин. Після колосіння (викидання волоті) у злакових або бутонізації у бобових та інших двосім'ядольних у рослинах прискорюється процес нагромадження сухої речовини, яка містить азотисті сполуки, клітковину, крохмаль, цукри, жир, фосфор, калій, кальцій та інші макро- і мікроелементи. В сухій речовині збільшується вміст клітковини (полісахариду), яка разом з лігніном, кутином і суберином спричинює загрубіння рослин. У деяких рослин воно відбувається відразу після настання генеративного періоду, наприклад у багаторічних і однорічних злакових трав, у зернових хлібів — пшениці, жита, ячменю, а також у проса, чумизи, могару та ін.
У бобових культур загрубіння відбувається повільніше. Їх добре поїдають тварини і у фазі цвітіння. Ще пізніше грубнуть однорічні бобові трави — вика яра, озима, паннонська, серадела, які добре поїдаються і після цвітіння, у фазі утворення бобиків. Винятком серед бобових кормових трав є буркун, який після бутонізації грубне майже так само, як і злакові трави. Довго не грубнуть зернобобові — горох, чина, боби, люпин, бо нагромадження клітковини у них починається після наливання бобів. Наприклад, рослини гороху у молочній стиглості зерна мають клітковини всього 12 — 14 %, тоді як злакові уже у фазі наливання зерна містять її понад 22 %. Досить довго не грубне кукурудза, вона добре поїдається тваринами до фази молочної стиглості качанів. Клітковини в ній у цей період небагато — 18 — 20 %. У фазі молочно-воскової стиглості зерна різко знижується перетравність сухої речовини у злакових, у тому числі в кукурудзи і сорго.
Достигання зерна супроводжується різким зниженням обводненості рослин, підсиханням листків і стебел. Рослини набувають солом'яного, коричневого і бурого кольору. Однак у просовидних злаків стебла й листя залишаються зеленими навіть при повному достиганні зерна (сорго, деякі гібриди кукурудзи, могар, чумиза, просо), що дає змогу господарству отримувати не тільки зерно, а й доброї якості і поживності грубі корми — стебла для заготівлі силосу, солому.
У дворічних коренеплідних рослин (буряків, брукви, моркви, турнепсу, цикорію, стеблоплідної капусти кормової) в перший рік вегетації формуються коренеплоди. На другому році ріст і розвиток відбуваються так само, як і в інших двосім'ядольних рослин. У багаторічних трав цей цикл повторюється 3 — 10 і більше років.
Слід відмітити також явище ремонтантності — безперервний розвиток у деяких культур (гречка, рицина, арахіс та ін.).
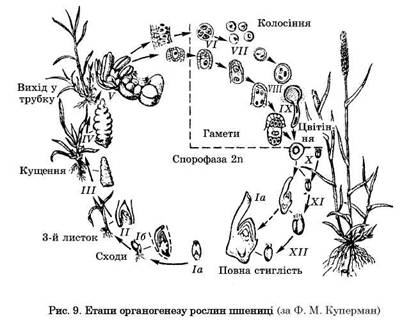
Етапи органогенезу. Розвиток органів пагона й суцвіття — безперервний процес. Зовнішні видимі зміни під час росту й розвитку рослин, які фіксуються у фазах вегетації, супроводжуються відповідними поступовими біохімічними, фізіологічними, гістологічними і на їх основі морфологічними змінами органів рослини, які формуються з меристеми пагонів. Розрізняють внутрішньобрунькову (ембріональну) і позабрунькову (постембріональну), вегетативну й генеративну фази, а також малий цикл розвитку — від розпукування бруньки до плодоношення.
Виділяють пагони з повним (закінченим) і перерваним (незакін-ченим) циклами розвитку. Повний цикл розвитку генеративного пагона може бути циклічним, якщо розвиток триває кілька років, дициклічним, коли розвиток пагонів завершується протягом двох років, і моноциклічним, якщо він відбувається за один вегетаційний період.
В органогенезі кожного пагона або кожної рослини виділяють 12 основних етапів:
I — виникнення на зародку апікальної ділянки меристеми конуса наростання головного пагона, формування зародкової бруньки.
На цьому етапі відбуваються процеси диференціації конуса наростання на різні тканини;
II — формується вегетативна сфера: вузли із зачатками листя та
міжвузлів;
III — формування головної генеративної осі пагона (осі початкового суцвіття);
IV — спостерігається гілкування осі генеративних пагонів, листя зазнає різних модифікацій і стає покривним — брактеями, приквіт никами або листковими обгортками, які зазвичай значно менші за справжні середні листки і часто редукуються;
V — відбуваються формування і диференціація квіток, заклада ються тичинки, приймочки та покривні органи квітки;
VI — це етап мікро- й макроспорогенезу. На цьому етапі у пилкових гніздах (мікроспорангіях тичинок, мікроспорофіл) з кожної материнської клітини пилку (мікроспороцита) в результаті мейозу
утворюються 4 мікроспори (тетрада) з гаплоїдним набором хромосом
у кожній. У покритонасінних мікроспорогенез відбувається здебі
льшого дещо раніше макроспорогенезу;
VII — формування чоловічого і жіночого гаметофітів. На цьому
етапі органогенезу дозрівають пилкові зерна і зародкові мішки, за
кінчується гаметогенез. За оптимальних умов на сьомому етапі ор
ганогенезу відбувається підготовчий процес до утворення основних
груп спеціалізованих клітин жіночого гаметофіту. Цей етап у пше
ниці і ячменю збігається з виколошуванням, у вівса й качана куку
рудзи — з викиданням волоті, в гороху і бобів — з розкриванням
квіток. Він зазвичай не супроводжується помітними змінами в бу
дові пилкових зерен і зародкових мішків; у яйцеклітині утворюється
багато дрібних крохмальних зерен;
VIII — фаза виколошування у злаків, бутонізації у бобових та
інших, тичинки і приймочки часто ще недозрілі, але квітки вже на
бувають вигляду типових і сортових ознак, характерного для при
цвітника забарвлення. У деяких видів (ячменю, бобових) важко
розрізнити восьмий і дев'ятий етапи, коли відбуваються процеси
запліднення, оскільки вони іноді самозапилюються до виколошу
вання і розкриття квіток;
IX — цвітіння, запліднення і утворення зиготи;
X — формується насіння, плоди розвиваються й інтенсивно рос
туть, досягаючи характерних розмірів для виду чи сорту;
XI — нагромадження поживних речовин у плодах і насінні, тобто
наливання зерна;
XII — поживні речовини перетворюються на запасні. При цьому
різко зменшується вміст води в плодах і насінні і вони повністю до
стигають. У фотосинтезуючих органах вже немає хлорофілу. Хімічні
процеси, які відбуваються при достиганні плодів, мають видову спе
цифіку: триває синтез білків, швидко розщеплюється крохмаль,
вміст цукрів залишається постійним або збільшується, вміст кислот
зменшується.
Дванадцятим етапом завершується повний цикл розвитку рослини, пагона, стебла. В однорічних рослин пагони або бруньки відновлення відмирають разом з рослиною (цим завершується великий цикл розвитку), у багаторічних рослин відмирають лише пагони, які відплодоносили, або їх квітконосна частина і зберігаються життєздатними бруньки або пагони відновлення, з яких наступного року розвиваються вегетативні й генеративні пагони. Етапи органогенезу (розвитку органів) пшениці зображені на рис. 9.
Життєвий цикл рослин можна поділити на кілька етапів (періодів): первинного спокою — починається від дозрівання і триває до проростання насіння; ювенільний (юнацький) — перша половина вегетаційного періоду до появи генеративних органів; генеративний — починається від початку цвітіння і плодоношення і триває до до-
зрівання насіння. У багаторічних трав ці цикли повторюються протягом 2 — 10 і більше років, але на другий і в наступні роки вони починаються спокоєм бруньок, які при проростанні перетворюються на пагін-сисунець. Після цього настає ювенільний період і т. д.
Якщо висівають свіжозібране насіння, період його первинного спокою триває від 8 - 40 до 200 - 210 днів.
Ювенільний період у багаторічних трав внаслідок перебування в рік сівби під покривною культурою, а в наступні роки під покривом інших трав може тривати від одного до кількох років. Затінені рослини розвиваються повільно, і лише при поліпшенні світлового режиму після укосів та внаслідок зрідження або випадання з травостою якогось компонента рослини з ювенільного переходять в генеративний період. Це має велике значення на багаторічних травостоях сіножатей і пасовищ. Завдяки цьому підтримується необхідна густота стеблостою агрофітоценозу.
Поділ рослин за тривалістю періоду вегетації. Сорти і гібриди польових культур умовно можна поділити на три основні великі групи: скоростиглі, середньостиглі, пізньостиглі. Між ними є перехідні групи. Наприклад, у кукурудзи розрізняють надранні — ульт-раскоростиглі, ранньостиглі, середньоранні, середньостиглі, середньопізні, пізньостиглі, ультрапізньостиглі сорти й гібриди. Так, різні гібриди та сорти кукурудзи достигають за 70 — 80 і за 130 — 160 днів і мають відповідно неоднакову продуктивність. Однак, коли ранньостиглі гібриди кукурудзи, рослини яких мають меншу вегетативну масу, висіяти густіше, то за короткий період вегетації можна мати майже такий самий врожай зерна, як і при висіванні пізньостиглих гідридів. При цьому забезпечують необхідний рівень живлення рослин. Тому ранньостиглі гібриди кукурудзи в Лісостепу і на Поліссі слід висівати з густотою 90- 100, а на силос 110- 130 тис. рослин на 1 га і отримувати по 90 — 100 ц/га зерна, 500 — 600 ц/га силосної маси.
Багаторічні трави поділяють на скоростиглі, які зацвітають наприкінці травня — на початку червня і дають стигле насіння в першій половині літа (еспарцет і грястиця збірна); середньостиглі — цвітуть у середині червня і достигають наприкінці липня — на початку серпня (люцерна посівна і жовта, вівсяниця лучна); пізньостиглі — цвітуть у другій половині червня, достигають у серпні. Цей поділ певною мірою також умовний, оскільки одні й ті самі види, як уже зазначалося, можуть мати ранні й пізні сорти, наприклад, грястиця збірна.
Поєднання різних за строками дозрівання культур на великих площах посіву дає можливість зменшити втрати врожаю від перестоювання, дощів, обсипання тощо.